Details of DPV and References
DPV NO: 51 June 1971
Family: Caulimoviridae
Genus: Caulimovirus
Species: Dahlia mosaic virus | Acronym: DMV
Dahlia mosaic virus
A. A. Brunt Glasshouse Crops Research Institute, Littlehampton, Sussex, England
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
Described by Brandenburg (1928), Brierley & Smith (1950) and Brunt (1971).
- Selected synonyms
- Dahlia
virus 1 (Rev. appl. Mycol. 17: 52) - Marmor dahliae (Rev. appl. Mycol. 18: 229)
- Dahliavirus maculans (Rev. appl. Mycol. 38: 677)
A virus with isometric particles c. 50 nm in diameter, probably containing DNA. Transmissible by inoculation of sap, and by 13 aphid species. It occurs naturally only in dahlias, but can be transmitted experimentally to other species. World-wide distribution.
Main Diseases
Causes chlorotic vein-banding (Fig. 1) followed by general mosaic and leaf distortion in many dahlia cultivars, and stunting of the most sensitive. Other cultivars are less seriously affected, and some remain almost symptomless.
Geographical Distribution
World-wide.
Host Range and Symptomatology
Natural infection found only in Dahlia species, but the virus can infect 11 other members of the Compositae, and 13 species in the Solanaceae, Chenopodiaceae and Amaranthaceae (Brunt, 1971). Transmissible by inoculation of sap.
-
Diagnostic species
- Verbesina encelioides.
Chlorotic or semi-necrotic circular local lesions 2-4 mm in diameter after 10-15 days (Fig. 2), especially in short day conditions, soon followed by chlorotic vein-banding (Fig. 3), general chlorosis and slight distortion of systemically infected leaves. - Dahlia pinnata. Infected readily by aphids, but with difficulty by mechanical
inoculation of sap; systemic chlorotic vein-banding at least 3 weeks after inoculation,
followed by general chlorosis and stunting.
- Ageratum conyzoides cv. Blue Ball. Chlorotic local lesions 2-4 mm in diameter after 10-20 days, followed by systemic chlorotic vein-banding and chlorosis.
- Zinnia elegans. Inoculated leaves usually symptomless; systemically infected leaves slightly distorted with inconspicuous and transient chlorosis after 14-21 days. Plants stunted.
- Amaranthus caudatus. Few chlorotic local lesions 1-2 mm in diameter after 10-14 days, followed by inconspicuous systemic mottling.
- Chenopodium capitatum. No local symptoms, but conspicuous systemic chlorotic vein- banding and chlorosis after 14-21 days.
- Ageratum conyzoides cv. Blue Ball. Chlorotic local lesions 2-4 mm in diameter after 10-20 days, followed by systemic chlorotic vein-banding and chlorosis.
-
Propagation species
- V. encelioides
is suitable for maintaining the virus, and is the best source of virus for purification.Assay species
- No reliable local lesion host, but may be assayed by recording the proportion of V. encelioides plants becoming infected.
Strains
Most isolates seem similar, but Brierley (1951) recorded a strain that was isolated with difficulty and gave relatively mild symptoms in Dahlia pinnata and Verbesina encelioides.
Transmission by Vectors
Transmissible by 13 aphid species, notably Aphis fabae, Myzus persicae and Macrosiphum euphorbiae (Heinze, 1951, 1952; Brierley & Smith, 1950). All instars transmit, but virus does not pass through the egg. Virus can be acquired in 1 min and inoculated in 5 min. There is no latent period. The virus can be retained both by feeding and by starved M. persicae for c. 3 h (Brierley & Smith, 1950), and it thus differs from typical ‘non-persistent’ viruses.
Transmission through Seed
None found (Brierley, 1933).
Transmission by Dodder
None recorded.
Serology
The virus is a fairly good immunogen when sufficiently concentrated (c. 0.1 mg/ml). Antisera react well in gel-diffusion tests to produce a single curved band of precipitate close to the antigen depôt within 48 h; the precipitin ring test is also useful, but microprecipitin and conventional precipitin tube tests are often unsatisfactory. Virus occurs in crude sap in insufficient amounts to be detected serologically. In gel immuno-electrophoresis in 0.8% Ionagar in 0.033 M phosphate buffer at pH 7.6 the virus moves slowly towards the anode at a rate of 5.3 x 10-7 cm2 sec-1 volt-1 (Brunt, 1971).
Relationships
The virus is serologically related both to cauliflower mosaic virus (Brunt, 1966) and to carnation etched ring virus (Hollings & Stone, 1969), but not to 19 other smaller isometric viruses (Brunt, 1966). Antisera to dahlia mosaic and cauliflower mosaic viruses differ little in homologous and heterologous titres in agar gel (0.8%) double diffusion tests. Although so closely related, neither virus infects the natural hosts of the other.
Stability in Sap
In Verbesina encelioides sap, the thermal inactivation point (10 min) is 75-80°C and the dilution end-point up to 1/3,000; infectivity is retained for 2-4 days at 18°C and for 16-32 days at 2°C (Brierley & Smith, 1950; Brunt, 1971). Sap from Dahlia pinnata and many other members of the Compositae rapidly loses infectivity unless a reducing agent (0.05 M thioglycollic acid, cysteine or ascorbic acid) or 0.01 M sodium diethyldithiocarbamate is present; such extracts remain infective for up to 48 h at 18°C. Mechanical transmission from dahlia to other hosts is also greatly aided by preparing extracts with liquid nitrogen (Lawson & Taconis, 1965).
Purification
Even in Verbesina encelioides, the best source, the virus occurs either in low concentration or is not readily released from intracellular inclusions; the use of recently infected leaves having a relatively high virus content seems more important than the type, molarity and pH of extractant. The following procedure gives yields of c. 0.25-1.0 mg of virus per kg of leaf tissue extracted. Triturate leaves in 0.05 M phosphate buffer containing either 0.3% thioglycollic acid or 0.01 M ascorbic acid at pH 7.0-8.0 (1 g leaf: 2 ml extractant), express the sap through cloth and add n-butanol to 8.5% (v/v). Store the mixture at 2°C for 10-14 days, then subject to one or more cycles of differential centrifugation (12,000 g for 15 min; 78,500 g for 45-60 min). Separate virus from remaining plant materials by centrifuging for 2 h at 53,500 g in 10-40% linear sucrose gradients.
Properties of Particles
Sedimentation coefficient (s20,w) at infinite dilution: 254 S. No accessory viral components found by analytical centrifugation.
A260/A280: 1.5; A260/A240: 1.16.
Particle Structure
Particles mounted in neutral 2% potassium phosphotungstate are isometric, c. 48-50 nm in diameter and similar to those of cauliflower mosaic virus (Day & Venables, 1960). Most of the particles mounted in saturated uranyl acetate or uranyl formate are slightly swollen or disrupted, even after fixation in 3% glutaraldehyde; surviving particles, however, show some surface structure and apparently have a protein shell c. 14-15 nm thick enclosing a hollow core c. 20 nm in diameter (Fig. 6). The particles are well preserved but contrast poorly in 2% ammonium molybdate.
Particle Composition
Nucleic acid: About 16% of particle weight (Brunt, 1971); like cauliflower mosaic virus (Shepherd, Bruening & Wakeman, 1970), probably DNA (R. J. Shepherd, unpublished data; Brunt, 1971).
Protein: About 84% of particle weight, probably only one kind of protein.
Relations with Cells and Tissues
Dahlia and all systemically infected hosts contain spherical or ellipsoidal intracellular inclusions (Goldstein, 1927) very similar to those induced by cauliflower mosaic virus (Rubio-Huertos, 1956). They occur in the cytoplasm of most living tissues, including immature tracheids, and especially in epidermal (Fig. 4), palisade and spongy parenchyma cells. Mature cells usually contain one inclusion 5-10 µm in diameter, but undifferentiated cells often contain several smaller inclusions. Inclusions frequently occur near, but never within, nuclei and are often associated with dictyosomes or Golgi cisternae (Kitajima, Lauritis & Swift, 1969). Inclusions in Verbesina encelioides (Brunt, 1969), dahlia (Kitajima & Lauritis, 1969) and Zinnia elegans (Kitajima et al., 1969), like those induced by cauliflower mosaic virus in members of the Cruciferae (Fujisawa et al., 1967), lack a limiting membrane, are usually surrounded by numerous ribosomes, and consist of an amorphous, vacuolated matrix containing numerous apparently hollow virus particles 40-45 nm in diameter (Fig. 5). Particles also sometimes occur freely in cytoplasm, in enlarged plasmodesmata (Kitajima & Lauritis, 1969) and within phloem sieve tubes and tracheids.
Notes
A virus described in France as dahlia mosaic (Cornuet, Martin & Limasset, 1950; Martin, 1954) was reported to have particles only 30 nm in diameter (Lépine et al., 1951), and therefore presumably differs from the virus described here.
Dahlia mosaic virus is prevalent because it is transmitted efficiently by several aphid species common on dahlias, and cultivars that produce only inconspicuous symptoms are often unsuspected and effective reservoirs of infection. Tobacco streak, tobacco rattle, tomato spotted wilt and cucumber mosaic viruses also infect dahlias; unlike dahlia mosaic, however, each of these viruses is readily isolated and identified serologically and/or by the reactions of Nicotiana glutinosa, N. tabacum, Chenopodium quinoa, C. amaranticolor and Vigna sinensis.
Dahlia mosaic and cauliflower mosaic viruses share many common properties and are closely related serologically, but have distinct host ranges. Therefore, they should still be regarded as distinct viruses which, with several others, form a small group. Carnation etched ring (Hollings & Stone, 1969), cassava vein mosaic (Kitajima & Costa, 1966) and an undescribed virus from Mirabilis nyctaginea (E. W. Kitajima & A. A. Brunt, unpublished) are probably members of this group. Carrot mottle virus (Murant et al., 1969) is not, because although it has particles 50 nm in diameter and occurs within plants in low concentration, it persists in its aphid vector for several days and has particles that contain RNA and, possibly, lipid.
Figures

Chlorotic vein-banding symptoms in naturally infected dahlia.

Chlorotic local lesions in Verbesina encelioides.

Systemic chlorotic vein-banding symptoms in V. encelioides.

Intracellular inclusions in leaf epidermal cells of Chenopodium quinoa (stained with 0.05% trypan blue). Bar represents 10 µm.

Intracellular inclusion in sectioned leaf cell of Zinnia elegans. Bar represents 500 nm. (Photo courtesy E. Kitajima; republished from Virology with the permission of The Academic Press Inc.)
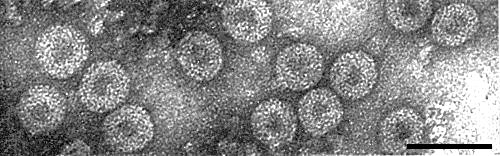
Virus particles from purified preparation mounted in saturated uranyl formate. Bar represents 100 nm.
References list for DPV: Dahlia mosaic virus (51)
- Brandenburg, Forschn Geb. Pflkrankh., Berl. 5: 39, 1928.
- Brierley, Contr. Boyce Thompson Inst. Pl. Res. 5: 235, 1933.
- Brierley, Pl. Dis. Reptr 35: 405, 1951.
- Brierley & Smith, Pl. Dis. Reptr 34: 363, 1950.
- Brunt, Virology 28: 778, 1966.
- Brunt, Rep. Glasshouse Crops Res. Inst., 1968: 104, 1969.
- Brunt, Ann. appl. Biol. 67: 357, 1971.
- Cornuet, Martin & Limasset, C. r. hebd. Séanc. Acad. Sci., Paris 231: 913, 1950.
- Day & Venables, Virology 11: 502, 1960.
- Fujisawa, Rubio-Huertos, Matsui & Yamaguchi, Phytopathology 57: 1130, 1967.
- Goldstein, Bull. Torrey bot. Club 54: 285, 1927.
- Heinze, Mitt. biol. Reichsanst. Ld-u. Forstw. 71: 126 pp., 1951.
- Heinze, Z. Pflkrankh. PflPath. PflSchutz 59: 3, 1952.
- Hollings & Stone, Rep. Glasshouse Crops Res. Inst., 1968: 102, 1969.
- Kitajima & Costa, Bragantia 25: 211, 1966.
- Kitajima & Lauritis, Virology 37: 681, 1969.
- Kitajima, Lauritis & Swift, Virology 39: 240, 1969.
- Lawson & Taconis, Phytopathology 55: 715, 1965.
- Lépine, Croissant, Cornuet & Martin, Annls Inst. Pasteur, Paris 80: 669, 1951.
- Martin, Annls Épiphyt. 5: 53, 1954.
- Murant, Goold, Roberts & Cathro, J. gen. Virol. 4: 329, 1969.
- Rubio-Huertos, Phytopathology 46: 553, 1956.
- Shepherd, Bruening & Wakeman, Virology 41: 339, 1970.