Details of DPV and References
DPV NO: 64 October 1971
Family: Unallocated ssRNA+ viruses
Genus: Sobemovirus
Species: Sowbane mosaic virus | Acronym: SoMV
Sowbane mosaic virus
C. I. Kado Department of Plant Pathology, University of California, Davis, California, USA
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
Described by
Silva, Meneghini & Lopes de Souza Santos (1958)
and
Bennett & Costa (1961).
Selected synonyms
- Chenopodium mosaic virus (Bragantia 17: 167, 1958)
- Apple latent virus 2 (Rev. appl. Mycol. 46: 3022; 46: 3705)
- Chenopodium star mottle virus (Rev. appl. Mycol. 46: 12m)
- Apple latent virus 2 (Rev. appl. Mycol. 46: 3022; 46: 3705)
-
An RNA-containing virus with isometric particles 26 nm in diameter, transmissible by inoculation with sap and possibly by various insects. Transmitted through seeds of several Chenopodium species. Widespread distribution.
Main Diseases
Causes chlorotic mottling of Chenopodium spp., many of which are employed as experimental assay hosts for other plant viruses. Test plants such as C. amaranticolor, C. murale and C. quinoa may carry the virus symptomlessly.
Geographical Distribution
Widespread; reported from Australia, Europe, Japan, North and South America, and South Africa.
Host Range and Symptomatology
Natural hosts are species of the Chenopodiaceae. A strain isolated from grapevines infects plants in other families experimentally (Bercks & Querfurth, 1969).
-
Diagnostic species
- Chenopodium amaranticolor.
Plants inoculated mechanically show chlorotic local lesions (Fig. 3) followed by yellow flecking and star-shaped patterns (Fig. 1, Fig. 2). Plants infected through seed may or may not develop mottling and flecking symptoms. Those that do not may be induced to show symptoms by rubbing their leaves with the finger moistened with water or buffer alone (Dias & Waterworth, 1967; Engelbrecht & Van Regenmortel, 1968). Symptoms in C. quinoa and C. murale are similar to those in C. amaranticolor. - Beta vulgaris (red beet). Very mild systemic mosaic.
-
Propagation species
- Chenopodium amaranticolor, C. quinoa
or C. murale are suitable for maintaining cultures. - Spinacia oleracea (spinach) is a good source of virus for purification.
-
Assay species
- Chenopodium amaranticolor
or C. quinoa give local lesions, but their suitability as assay hosts has not been tested.
Strains
All isolates are serologically indistinguishable. They include: apple latent virus 2 of Kirkpatrick et al. (1965) and Bancroft & Tolin (1967); carnation 698 isolate of Hollings & Stone (1967); chenopodium mosaic virus of Silva et al. (1958); chenopodium star mottle virus of Kado (1966, 1967); Chenopodium trigonon (fish weed) virus of Teakle (1968); and an isolate from grapevine (Bercks & Querfurth, 1969). All seem very similar except the grapevine isolate which differs from the others because it infects experimentally species of the Amaranthaceae, Cucurbitaceae, Leguminosae and Solanaceae.
Transmission by Vectors
Liriomyza langei (leafminer fly), Circulifer tenellus (beet leafhopper), Halticus citri (fleahopper) and Myzus persicae (green peach aphid) are reported as probable vectors (Bennett & Costa, 1961). However, Dias & Waterworth (1967) could not confirm that M. persicae transmits the virus.
Transmission through Seed
Occurs in up to 83%, of progeny seedlings of infected plants, depending on the seed sample (Bennett & Costa, 1961; Dias & Waterworth, 1967; Kado, 1967). Plants infected through seed may show few or no symptoms.
Transmission by Dodder
No transmission was obtained with Cuscuta californica which apparently is immune to infection (Bennett & Costa, 1961).
Serology
The virus is strongly immunogenic. Double-diffusion tests in buffered 1% agar or agarose gels, precipitin tests in tubes, and immuno-electrophoresis in 1% agar or agarose gel may be used. A single precipitin band usually forms in gel double-diffusion and immuno-electrophoresis tests. In tube precipitin tests, the precipitates are granular.
Relationships
No serological relationship has been detected to the following viruses: alfalfa mosaic, arabis mosaic, bean pod mottle, broad bean mottle, cowpea mosaic, cucumber mosaic, cucumber mosaic strain Y, grapevine fanleaf, peach yellow bud mosaic, southern bean mosaic, squash mosaic, tobacco necrosis, tobacco ringspot, tomato aspermy, tomato bushy stunt, tomato spotted wilt, turnip yellow mosaic and wild cucumber mosaic.
Stability in Sap
In Chenopodium quinoa sap, the thermal inactivation point (10 min) is about 90°C, dilution end-point about 10-7, and infectivity is retained at 23°C for about 60 days. Infectivity survives freezing indefinitely.
Purification
Yields of 200-500 mg virus per kg inoculated leaf tissue are obtained. Either of two methods may be used:
1. Kado (1967). Grind frozen tissue in 0.05 M sodium ethylenediamine-tetraacetate, pH 7.5. Filter and add 0.5 vol of cold n-butanol, stir, and centrifuge to break emulsion. Remove upper butanol phase by aspiration and filter aqueous phase through glass wool. Concentrate by five cycles of high and low speed centrifugation, suspending the virus pellets in 0.1 M phosphate buffer, pH 7.0.
2. The butanol-chloroform method of Steere (1956) may be used.
Properties of Particles
Only one sedimenting component. Sedimentation coefficient at infinite dilution (s20, w): 104±2 S.
Molecular weight: 6.3±0.4 x 106 daltons (Kado & Black, 1968).
Diffusion coefficient (D20 x 10-7 cm2/sec): 1.29 (calculated: Kado, unpublished); 1.15 (Paul & Huth, 1970).
Isoelectric point: pH 4.4.
Partial specific volume (calculated): 0.70±0.05 cm3/g.
Absorbance at 260 nm (1 mg/ml, 1 cm light path): 4.9.
A260/A280 (corrected for light scatter): 1.49; A230/A260: 1.67.
Particle Structure
Particles are isometric, 26±2 nm in diameter (Fig. 4). Existing electron micrographs of particles stained with phosphotungstate or uranyl acetate do not resolve any morphological subunits (Fig. 4).
Particle Composition
RNA: M. Wt c. 1.3 x 106, about 20% of particle weight, probably single-stranded. Molar percentages of nucleotides: G26.0; A23.2; C27.6; U23.2.
Protein: Particles contain one type of protein subunit. The C-terminal amino acid is lysine, the N-terminus is blocked. Amino acid composition (moles %): ala 8.5; arg 4.6; asp 9.1; cys 1.1; glu 6.8; gly 9.1; his 1.7; ile 5.1; leu 6.8; lys 6.8; met 2.8; phe 2.3; pro 6.8; ser 8.0; thr 7.4; trp 1.7; tyr 4.0; val 7.4 (Kado, 1967). Amino acid analysis suggested that the M. Wt of the protein subunit is 1.92 x 104 (Kado, 1967) but electrophoresis in neutral sodium dodecyl sulphate/polyacrylamide gels indicates that the M. Wt is probably c. 3.1 x 104 (Hill, 1971).
Relations with Cells and Tissues
All tissues infected. Virus occurs in the cytoplasm as membrane-bound crystalline arrays and induces ‘pseudonucleoli’-dense, circular, granular regions (Milne, 1967).
Notes
The fact that seed of Chenopodium species may carry sowbane mosaic virus can present contamination problems when those species are used as hosts for other viruses, especially because infected progeny seedlings may show no symptoms. Care should be taken to use only seed collected from plants free from sowbane mosaic virus.
Figures

Chlorotic mottling in systemically infected leaf of Chenopodium amaranticolor.

Yellow flecking and star-shaped patterns in systemically infected leaf of C. amaranticolor.

Chlorotic local lesions in inoculated leaf of C. amaranticolor.
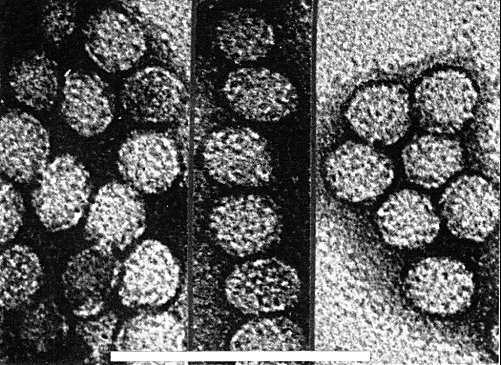
Virus particles from a purified preparation negatively stained with uranyl acetate, pH 4.2. Bar represents 100 nm.
References list for DPV: Sowbane mosaic virus (64)
- Bancroft & Tolin, Phytopathology 57: 693, 1967.
- Bennett & Costa, Phytopathology 51: 546, 1961.
- Bercks & Querfurth, Phytopath. Z. 66: 365, 1969.
- Dias & Waterworth, Can. J. Bot. 45: 1285, 1967.
- Engelbrecht & Van Regenmortel, Phytopath. Z. 63: 10, 1968.
- Hill, Doctoral Dissertation, Univ. California, Davis, 1971.
- Hollings & Stone, Rep. Glasshouse Crops Res. Inst. 1966: 91, 1967.
- Kado, Phytopathology 56: 884, 1966.
- Kado, Virology 31: 217, 1967.
- Kado & Black, Virology 36: 137, 1968.
- Kirkpatrick, Mink & Lindner, Phytopathology 55: 286, 1965.
- Milne, Virology 32: 589, 1967.
- Paul & Huth, Phytopath. Z. 69: 1, 1970.
- Silva, Meneghini & Lopes de Souza Santos, Bragantia 17: 167, 1958.
- Steere, Phytopathology 46: 60, 1956.
- Teakle, Aust. J. biol. Sci. 21: 649, 1968.