Details of DPV and References
DPV NO: 69 October 1971
Family: Tombusviridae
Genus: Tombusvirus
Species: Tomato bushy stunt virus | Acronym: TBSV
There is a more recent description of this virus: DPV 382
Tomato bushy stunt virus
G. P. Martelli Istituto di Patologia vegetale, Università di Bari, 70126 Bari, Italy
A. Quacquarelli Istituto di Patologia vegetale, Università di Bari, 70126 Bari, Italy
M. Russo Istituto di Patologia vegetale, Università di Bari, 70126 Bari, Italy
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
Described by
Smith (1935),
Ainsworth (1936) and
Bawden & Pirie (1938).
Selected synonyms
- Lycopersicum
virus 4 (Rev. appl. Mycol. 17: 52) - Marmor dodecahedron (Rev. appl. Mycol. 28: 514)
- Lycopersicumvirus dodecahedron (Rev. appl. Mycol. 38: 677)
- Tomatenzwergbusch-Virus (Rev. appl. Mycol. 37: 740)
- Lycopersicumvirus dodecahedron (Rev. appl. Mycol. 38: 677)
-
An RNA-containing virus with isometric particles c. 30 nm in diameter. Stable in crude plant extracts and readily transmissible by inoculation of sap. Has a wide experimental host range; in nature infects a few dicotyledonous species in widely separated families. No vector known.
Main Diseases
Causes stunting and bushy growth of tomato, mottle, severe leaf deformation, reduced growth and sterility of artichoke (Fig. 1) (Quacquarelli & Martelli, 1966), distortion and yellow stellate spots in Pelargonium leaves (Fig. 2) (Pape, 1927; Hollings, 1962), asteroid mosaic of Petunia (Fig. 3) (Lovisolo, 1957), fruit-pitting, veinal necrosis and stunting of cherry (Allen & Davidson, 1966), and chlorotic spots and rings in carnation leaves (Hollings, Stone & Bouttell, 1970). Also obtained from grapevine (Bercks, 1967).
Geographical Distribution
Several European countries; North (Allen & Davidson, 1966) and South America (Pontis, Gracia & Feldman, 1968).
Host Range and Symptomatology
Transmitted experimentally to over 120 species in more than 20 families (Schmelzer, 1958; Hollings, 1962; Hollings & Stone, 1965; Hollings et al., 1970). Most hosts are not invaded systemically.
-
Diagnostic species
- Chenopodium amaranticolor.
Whitish local necrotic dots, sometimes with chlorotic haloes. Limited systemic infection with some strains. - Gomphrena globosa. Pale necrotic local lesions which enlarge to give
red-rimmed whitish rings or irregular spots
(Fig. 5).
Limited systemic infection
with some strains.
- Nicotiana clevelandii. Chlorotic or necrotic local lesions which may develop into necrotic rings. Severe chlorotic systemic mottling, malformation and stunting, killing some plants.
- Nicotiana glutinosa. Brown necrotic local lesions in 2-3 days, darker at the edge than at the centre, often enlarging to 5 mm. Rarely, a few systemic lesions develop.
- Ocimum basilicum (basil). Dark brown necrotic local lesions with lighter centres (Fig. 6). Not systemic.
- Nicotiana clevelandii. Chlorotic or necrotic local lesions which may develop into necrotic rings. Severe chlorotic systemic mottling, malformation and stunting, killing some plants.
- Propagation species
- Nicotiana clevelandii.
Alternatively, Datura stramonium, Petunia hybrida or Chenopodium quinoa can be used. Some strains have been purified from natural hosts, e.g. Petunia (Lovisolo et al., 1964), artichoke (Quacquarelli. Martelli & Russo, 1966) and cherry (Allen, 1968).Assay species
- Chenopodium amaranticolor, Datura stramonium
and Ocimum basilicum are useful local lesion hosts. - Nicotiana glutinosa, Celosia argentea and Phaseolus vulgaris are also satisfactory for some strains.
Strains
The following major variants are described:
Type strain (Smith, 1935). Causes ring-like or necrotic local lesions, severe stunting, yellowing and distortion of the leaves of Lycopersicon esculentum. In Datura stramonium it causes local circular dendritic yellow spots, followed by systemic crinkling and green-yellow mottling.
Pelargonium strain (pelargonium leaf curl ) (Pape, 1927; Hollings, 1962). Induces pale brown local necrotic spots and stellate lesions in Phaseolus vulgaris, followed by systemic mottle. In Datura stramonium, local chlorotic lesions, becoming necrotic, followed by systemic symptoms. Infects Chenopodium amaranticolor systemically.
Petunia strain (petunia asteroid mosaic) (Lovisolo, 1957). Unlike other strains, infects Petunia systemically. The grapevine isolate behaves in this way (Bercks, 1967).
Carnation strain (carnation Italian ringspot) (Hollings & Stone, 1965; Hollings et al., 1970). Produces local and systemic infection in Chenopodium amaranticolor and C. quinoa.
Artichoke strain (artichoke mottled crinkle) (Martelli, 1965; Quacquarelli & Martelli, 1966). Induces pin-point local lesions in Phaseolus vulgaris cv. Bountiful, small necrotic lesions but no systemic infection in Datura stramonium and small lesions in Nicotiana glutinosa.
Cherry strain (Allen & Davidson, 1966). Induces mosaic, chlorotic line pattern, rings and green blisters in Datura stramonium and systemic top necrosis in Chenopodium amaranticolor.
Minor variants occur within most of the major strains e.g. the BS-3 and BS-9 variants of the type strain (Steere, 1953; De Fremery & Knight, 1955). These are distinguished by the symptoms they cause in hosts such as Datura stramonium, Chenopodium amaranticolor, Nicotiana glutinosa and N. clevelandii.
Transmission by Vectors
No insect or mite vector (Orlob, 1968) is known. Lovisolo, Bode & Volk (1965) and Lovisolo (1966) showed that the Petunia strain is transmitted through the soil and suggested that a fungus vector might be involved. However, trials with Olpidium brassicae zoospores gave either negative or inconclusive results (Teakle & Gold, 1963; Campbell, 1968; Allen & Davidson, 1966).
Transmission through Seed
None found (Hollings, 1962; Lovisolo et al., 1965) but the virus is present in the pollen (Allen & Davidson, 1966).
Transmission by Dodder
Cuscuta campestris can transmit the type strain (Johnson, 1941) and becomes infected by the Pelargonium strain (Hollings, 1962).
Serology
Antisera with precipitin titres up to 1/16,834 (M. Hollings, pers. comm.) can be produced by intravenous and intramuscular injections. Tube-precipitin (granular precipitates), or gel-diffusion and intra-gel absorption tests (single precipitin bands) can be used for comparing strains of the virus. The strains differ widely in immunoelectrophoretic behaviour (Bercks & Lovisolo, 1965; Martelli & Quacquarelli, 1966; Hollings et al., 1970) but consist of only one antigenic component.
Relationships
The carnation and type strains do not react with antiserum to each other but each reacts with antiserum to the Pelargonium strain (Hollings & Stone, 1965). The Petunia and cherry strains are more closely related to the Pelargonium than to the type strain (Wetter & Luisoni, 1969; Allen, 1968) whereas the reverse seems to be true with the artichoke strain (Martelli & Quacquarelli, 1966).
Stability in Sap
The infectivity dilution end-point is 10-2-10-6 depending on the strain and host used. Thermal inactivation point is between 80 and 90°C. At 20°C, infectivity is retained for 4-5 weeks. The carnation strain was infective for 23 weeks at 2°C and other strains retained infectivity for at least 10 years when lyophilized (Hollings & Stone, 1970; Hollings et al., 1970).
Purification
Steere (1959) obtained satisfactory preparations by differential centrifugation of sap. Other methods include the following (Hollings, 1962). Mince leaves in 1.25 vol. 0.05 M phosphate buffer pH 7.8 containing 0.1% Na thioglycollate. Add n-butanol dropwise to 8.5% (v/v). Allow to stand for 24 hr at 2°C, then clarify the mixture by low speed centrifugation. After differential centrifugation of the supernatant fluid, resuspend the pellets in 0.03 M phosphate buffer.
The artichoke strain has been purified from water extracts of artichoke. Extracts clarified with magnesium-activated bentonite and/or chloroform-butanol yielded up to 100 mg virus per kg leaf tissue (Quacquarelli et al., 1966).
When centrifuged in density gradients a single opalescent band is usually formed. The Pelargonium and carnation strains, however, may yield up to four light-scattering zones (Hollings, 1962; Hollings et al., 1970).
Properties of Particles
Sedimentation coefficients (s20,w) for the different strains range between 131 and 140 S.
Molecular weight (daltons): 9.3 x 106 (Weber, Rosenbusch & Harrison, 1970).
Diffusion coefficient (D20 x 10-7 cm2/sec): 1.26 (Schachman & Williams, 1959).
Isoelectric point: pH 4.1 (McFarlane & Kekwick, 1938).
Partial specific volume: 0.71 cm3/g (Schachman & Williams, 1959).
Electrophoretic mobility: -4.96 x 105 and -4.65 x 105 cm2 sec-1 volt-1, for Petunia and type (BS-3) strains respectively, at pH 8.2 in veronal-HCl buffer of ionic strength 0.02 (Ambrosino et al., 1967). Zone electrophoretic Rø values (Van Regenmortel, 1968) are 0.17, 0.25 and 0.34 for Pelargonium, artichoke and type strains respectively (Quacquarelli et al., 1966).
Absorbance at 260 nm (1 mg/ml, 1 cm light path): 4.5 (Ambrosino et al., 1967).
A260/A280: 1.62 and 1.66 for Petunia and type (BS-3) strains respectively (Ambrosino et al., 1967).
Particle Structure
Particles are isometric, c. 30 nm in diameter (Fig. 4). In phosphotungstate a few particles are penetrated, perhaps because they are damaged. Preparations of some strains must be fixed with formaldehyde before mounting for electron microscopy. The protein shell of the particle contains 90 morphological units (each composed of two protein molecules, making 180 in all) located on the two-fold axes of a T = 3 icosahedral surface lattice (Finch, Klug & Leberman, 1970). Moreover, chemical and physical data (Weber et al., 1970; Butler, 1970) indicate that the particle contains no more than 180 molecules of the major protein present, thus excluding the possibility, which has been suggested, of an inner protein core. Butler (1970) suggested that molecules of one of two minor proteins are situated on the five-fold axes of the particles.
Particle Composition
RNA: M. Wt c. 1.5 x 106, 16-17% of particle weight, single-stranded. Molar percentages of nucleotides of type (BS-3) strain: G28.6; A26.3; C21.2; U26.3 (De Fremery & Knight, 1955); data for other strains are given by Quacquarelli et al. (1966), Ambrosino et al. (1967) and Tremaine (1970).
Protein: Subunits of the major protein in the particle have M. Wt of 3.8-4.2 x 104 (Weber et al., 1970; Michelin Lausarot et al., 1970; Butler, 1970) and contain 380-400 amino acid residues (Michelin-Lausarot et al., 1970). Butler (1970) reported that the particles contain a few molecules of protein with M. Wts of 2.8 x 104 and 8 x 104 but the origin of these proteins is not known. Amino acid composition of the total protein of the type (BS-3) strain (mg/100 mg): ala 5.9; arg 7.0; asp 11.6; cys 0.9; glu 6.8; gly 5.5; his 1.6; ile 3.9; leu 11.7; lys 4.2; met 0.9; phe 4.8; pro 3.6; ser 7.1; thr 10.2; tyr 3.6; val 10.1 (Michelin-Lausarot et al., 1970). Analyses of proteins of other strains e.g. BS-9 (De Fremery & Knight, 1955), cherry and type (Tremaine, 1970), BS-9 and Petunia (Michelin-Lausarot et al., 1970), show that they differ little in amino acid composition.
Relations with Cells and Tissues
Numerous virus particles occur in conducting and parenchyma tissues (Russo, Martelli & Quacquarelli, 1967, 1968). Particles of the artichoke strain occur in the cytoplasm, either scattered or arranged in crystals with a cubic close-packed structure (Fig. 7). They are sometimes extruded into the vacuole in vesicles (Fig. 8) (Russo et al., 1968). The Pelargonium strain behaves similarly and its particles are also associated with the nucleus (Martelli & Castellano, 1969).
Notes
Symptoms in some experimental hosts resemble those caused by tobacco necrosis virus but the latter differs in remaining localized in Nicotiana clevelandii and Datura stramonium.
Turnip crinkle virus has particles structurally very similar to those of tomato bushy stunt virus and, like it, is very stable in vitro. However, the viruses differ greatly in host range and symptomatology and seem serologically unrelated (Hollings & Stone, 1965).
Acknowledgements
Photographs 3 and 5 courtesy of Dr O. Lovisolo, Laboratorio di Fitovirologia applicata del CNR, Torino, Italy.
Figures

Artichoke infected with artichoke strain.

Chlorotic spots and distortion on leaf of pelargonium infected with the Pelargonium strain.
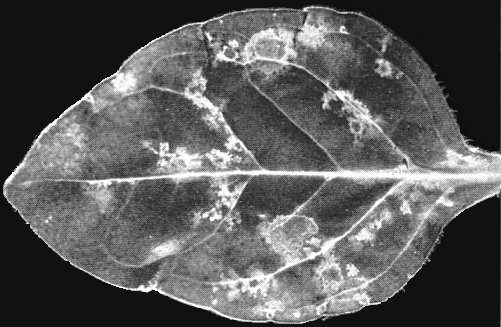
Petunia infected with the Petunia strain.

A purified preparation of the artichoke strain mounted in phosphotungstate. Bar represents 100 nm.

Local lesions in an inoculated leaf of Gomphrena globosa.

Local lesions in an inoculated leaf of Ocimum basilicum.

Thin section of an infected artichoke leaf showing a virus crystal in the cytoplasm. Bar represents 250 nm.

Thin section of an infected artichoke leaf showing a virus-containing vesicle protruding from the cytoplasm into the vacuole. Bar represents 250 nm.
References list for DPV: Tomato bushy stunt virus (69)
- Ainsworth, J. Minist. Agric. Fish. 43: 266, 1936.
- Allen, Can. J. Bot. 46: 229, 1968.
- Allen & Davidson, Can. J. Bot. 45: 2375, 1966.
- Ambrosino, Appiano, Rialdi, Papa, Redolfi & Carrara, Atti Accad. Sci., Torino 101: 301, 1967.
- Bawden & Pirie, Br. J. exp. Path. 19: 251, 1938.
- Bercks, Phytopath. Z. 60: 273, 1967.
- Bercks & Lovisolo, Phytopath. Z. 52: 96, 1965.
- Butler, J. molec. Biol. 52: 589, 1970.
- Campbell, Pl. Dis. Reptr 52: 379, 1968.
- De Fremery & Knight, J. biol. Chem. 214: 559, 1955.
- Finch, Klug & Leberman, J. molec. Biol. 50: 215, 1970.
- Hollings, Ann. appl. Biol. 50: 189, 1962.
- Hollings & Stone, Ann. appl. Biol. 56: 87, 1965.
- Hollings & Stone, Ann. appl. Biol. 65: 411, 1970.
- Hollings, Stone & Bouttell, Ann. appl. Biol. 65: 299, 1970.
- Johnson, Phytopathology 31: 649, 1941.
- Lovisolo, Boll. Staz.Patol. veg. Roma, S III 14: 103, 1957.
- Lovisolo, Atti Congr. Un. fitopat. Medit. 1: 574, 1966.
- Lovisolo, Bode & Volk, Phytopath. Z. 53: 323, 1965.
- Lovisolo, Ambrosino, Liberatori & Papa, Atti Accad. Sci., Torino 98: 391, 1964.
- Martelli, Phytopath. Mediterranea 4: 58, 1965.
- Martelli & Castellano, Virology 39: 610, 1969.
- Martelli & Quacquarelli, Atti Congr. Un. fitopat. Medit. 1: 195, 1966.
- McFarlane & Kekwick, Biochem. J. 32: 1607, 1938.
- Michelin-Lausarot, Ambrosino, Steere & Reichmann, Virology 41: 160, 1970.
- Orlob, Virology 35: 121, 1968.
- Pape, Gartenwelt, Berl. 31: 329, 1927.
- Pontis, Gracia & Feldman, Pl. Dis. Reptr 52: 676, 1968.
- Quacquarelli & Martelli, Atti Congr. Un. fitopat. Medit. 1: 168, 1966.
- Quacquarelli, Martelli & Russo, Atti Congr. Un. fitopat. Medit. 1: 163, 1966.
- Russo, Martelli & Quacquarelli, Virology 33: 555, 1967.
- Russo, Martelli & Quacquarelli, Virology 34: 679, 1968.
- Schachman & Williams, in The Viruses ed. F. M. Burnet & W. M. Stanley, Academic Press, New York, 1959.
- Schmelzer, Z. PflKrankh. PflPath. PflSchutz 65: 80, 1958.
- Smith, Ann. appl. Biol. 22: 731, 1935.
- Steere, Phytopathology 43: 485, 1953.
- Steere, Adv. Virus Res. 6: 1, 1959.
- Teakle & Gold, Virology 19: 310, 1963.
- Tremaine, Phytopathology 60: 454, 1970.
- Van Regenmortel, S. Afr. med. J. 42: 118, 1968.
- Weber, Rosenbusch & Harrison, Virology 41: 763, 1970.
- Wetter & Luisoni, Phytopath. Z. 65: 231, 1969.