Details of DPV and References
DPV NO: 91 June 1972
Family: Potyviridae
Genus: Potyvirus
Species: Parsnip mosaic virus | Acronym: ParMV
Parsnip mosaic virus
A. F. Murant Scottish Horticultural Research Institute, Invergowrie, Dundee, Scotland
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
- Described by Murant, Munthe & Goold (1970).
- A virus with flexuous filamentous particles about 730-760 x 14 nm, transmitted by aphids after brief acquisition feeds and by mechanical inoculation of sap. Infects several umbelliferous species and a few species in other families. Found in UK.
Main Diseases
Causes a mild mosaic disease of parsnip (Pastinaca sativa). It occurs commonly in parsnip crops in Britain, often in mixed infections with other viruses, such as parsnip yellow fleck and parsnip mottle (Murant & Goold, 1968), but its economic importance, if any, is unknown.
Geographical Distribution
Reported only from UK.
Host Range and Symptomatology
Infects several species in the Umbelliferae and also a few members of the Amaranthaceae, Chenopodiaceae and Scrophulariaceae. Transmissible by manual inoculation of sap.
- Diagnostic species
- Chenopodium amaranticolor
. Discrete necrotic local lesions 1-2 mm diameter in 6 days, later developing chlorotic margins (Fig. 1). Subsequently the inoculated leaves become chlorotic and absciss. No systemic infection. - C. quinoa. Irregular papery necrotic local lesions 1-2 mm diameter
in 6 days (Fig. 2). Subsequently the lesions coalesce and inoculated leaves
become yellow and absciss. No systemic infection.
- Anthriscus cerefolium (chervil). Fine systemic veinal necrosis visible on the youngest leaves after 8 days, followed by veinal chlorosis and distortion (Fig. 4). Plants are stunted but not usually killed.
- Coriandrum sativum (coriander). Severe systemic veinal necrosis and distortion of youngest leaves in 8-14 days (Fig. 3). Plants may die after 2-3 weeks.
- Pastinaca sativa (parsnip). Yellow systemic vein-banding in 8-10 days. Plants subsequently become almost symptomless.
- Anthriscus cerefolium (chervil). Fine systemic veinal necrosis visible on the youngest leaves after 8 days, followed by veinal chlorosis and distortion (Fig. 4). Plants are stunted but not usually killed.
- Propagation species
- The virus can be propagated in inoculated leaves of Chenopodium quinoa.
Isolates can be maintained in Coriandrum sativum.
- Assay species
- Chenopodium amaranticolor and C. quinoa are good local lesion hosts. Chervil and coriander are suitable test plants for experiments on transmission by aphids.
Strains
None reported.
Transmission by Vectors
Transmitted in the non-persistent manner by the aphids Cavariella aegopodii, C. theobaldi and Myzus persicae. Aphids acquired virus after 2-5 min feeding. Starving the aphids for 2-4 hr before the acquisition feed increased the efficiency of transmission (Murant et al., 1970).
Transmission through Seed
No information.
Transmission by Dodder
No information.
Serology
Antiserum with a titre of 1/256 was obtained from a rabbit given a single intramuscular injection with partially purified virus emulsified with Freund’s complete adjuvant (Murant et al., 1970). Ring precipitin or microprecipitin tests can be done using partially purified virus preparations from Chenopodium quinoa as antigen.
Relationships
In particle morphology and in its in vivo and in vitro properties, parsnip mosaic virus resembles members of the potato virus Y group (potyviruses). However, no serological relationship was detected to any of eleven viruses in this group (Murant et al., 1970), including two that infect umbelliferous plants, celery mosaic and clover yellow vein. In more recent work (Murant & Goold, 1972) no reaction was obtained with antisera to viruses from parsley and poison hemlock studied by Sutabutra & Campbell (1971).
Stability in Sap
In extracts of Chenopodium quinoa leaves, the virus lost infectivity after dilution to 10-3-10-4, or storage for 10 min at 55-58°C or 7-10 days at 18°C (Murant et al., 1970).
Purification
Murant et al. (1970) used the following procedure: homogenize Chenopodium quinoa inoculated leaves (100 g) in 200 ml 0.1 M borate, 0.01 M ethylene diamine tetra acetate, pH 8.0, containing 0.1% thioglycollic acid, and filter through cheesecloth. Emulsify the filtrate with an equal volume of chloroform and centrifuge at low speed. Concentrate by one cycle of differential centrifugation, dispersing the high speed pellets in borate/EDTA buffer, pH 8.0. Precipitate the virus by adjusting the pH to 5.0, then resuspend at pH 8.0 the pellets produced by low speed centrifugation. Remove phytoferritin-like particles of host origin by exclusion chromatography in a column of 2% agarose beads, having first dialysed the preparation against 0.15 M NaCl, 0.01 M EDTA, pH 8.0, the buffer used as the eluant for the column. Virus preparations obtained in this way are substantially free from host material but many particles are aggregated. Aggregation can be largely prevented by eliminating the pH 5.0 precipitation step.
Properties of Particles
Sedimentation coefficient, s20,w: 149 S.
Particle Structure
The particles are flexuous filaments 14 nm wide with a modal length of 736 nm in sap from Chenopodium quinoa containing phosphotungstate at pH 6.8 (Murant et al., 1970). In partially purified preparations (Fig. 5) the particles have a modal length of 755 nm.
Particle Composition
No information.
Relations with Cells and Tissues
Virus particles were rarely seen in ultrathin sections of systemically infected coriander leaf cells, but the cells contained abundant pinwheel and bundle inclusions (Fig. 6) resembling those described for other potyviruses (Murant & Roberts, 1971).
Notes
Several viruses with filamentous particles occur in umbelliferous plants. Celery mosaic virus was considered to be restricted to the Umbelliferae (Shepherd & Grogan, 1971) but there are several recent reports that some isolates thought to be celery mosaic virus can infect Chenopodium species. Such isolates were obtained from celery by Wolf (1969), from carrot by Iwaki & Komuro (1970) and from parsley and poison hemlock by Sutabutra & Campbell (1971). Only the last two viruses were shown to be serologically related to celery mosaic virus, though distinguishable from it. Parsnip mosaic virus does not react with antisera to these two viruses nor with antiserum to celery mosaic virus itself. Other viruses with filamentous particles that infect umbelliferous plants include carrot mosaic (Chod, 1965), celery latent (Brandes & Luisoni, 1966) and clover yellow vein viruses (Singh & López-Abella 1971). These, however, infect several non-umbelliferous species systemically, whereas parsnip mosaic virus does not.
There are similarities in host range between parsnip mosaic virus and a virus with filamentous particles from parsley (Férault et al., 1969) and another of unknown particle morphology from Heracleum sphondylium (Polák, 1966), but no serological comparisons have been reported.
Figures

Lesions in inoculated leaf of Chenopodium amaranticolor.

Lesions in inoculated leaf of C. quinoa.

Systemic veinal necrosis and distortion in coriander (Coriandrum sativum).

Systemic vein-yellowing and distortion in chervil (Anthriscus cerefolium).

Particles in a partially purified preparation mounted in 2% potassium phosphotungstate, pH 6.8. Bar represents 250 nm.
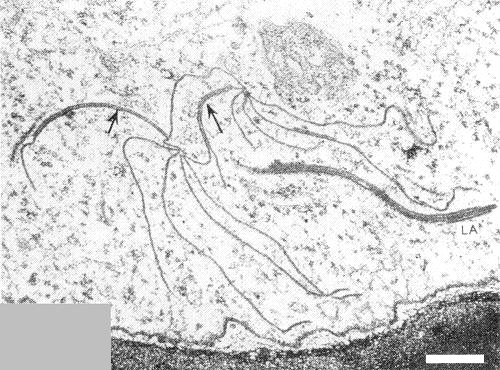
Section of systemically infected coriander leaf showing pinwheels linked (arrows) through the formation of laminated aggregates (LA). Bar represents 200 nm.
References list for DPV: Parsnip mosaic virus (91)
- Brandes & Luisoni, Phytopath. Z. 57: 277, 1966.
- Chod, Ochr. Rost. 3: 49, 1965.
- Férault, Lapierre, Spire & Bertrandy, Annls Phytopath. 1: 303, 1969.
- Iwaki & Komuro, Ann. phytopath. Soc. Japan 36: 36, 1970.
- Murant & Goold, Ann. appl. Biol. 62: 123, 1968.
- Murant & Goold, Rep. Scott. hort. Res. Inst. 1971 64, 1972.
- Murant, Munthe & Goold, Ann. appl. Biol. 65: 127, 1970.
- Murant & Roberts, J. gen. Virol. 10: 65, 1971.
- Polák, Biologia Pl. 8: 73, 1966.
- Shepherd & Grogan, CMI/AAB Descriptions of Plant Viruses 50, 4 pp., 1971.
- Singh & López-Abella, Phytopathology 61: 333, 1971.
- Sutabutra & Campbell, Pl. Dis. Reptr 55: 328, 1971.
- Wolf, Mber. dt. Akad. Wiss. Berl. 11: 293, 1969.