Details of DPV and References
DPV NO: 10 June 1970
Family: Caulimoviridae
Genus: Badnavirus
Species: Cacao swollen shoot virus | Acronym: CSSV
Cacao swollen shoot virus
A. A. Brunt Glasshouse Crops Research Institute, Littlehampton, Sussex, England
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
Described by Posnette (1947).
Selected synonyms
- Theobroma
virus 1 (Rev. appl Mycol. 27: 178) - Marmor theobromae (Rev. appl. Mycol. 27: 178)
- Theobromavirus inflans (Rev. appl. Mycol. 38: 677)
-
A virus of great economic importance in West Africa, with numerous symptomatologically distinct variants and a restricted host range; transmitted by 14 mealybug species, which retain it for a few hours. Can be mechanically transmitted to cacao beans and has bacilliform particles c. 121-130 x 28 nm.
Main Diseases
In Theobroma cacao, most strains induce conspicuous symptoms and reduce yield, but are non-lethal unless subsequent attack by capsids debilitates trees further and allows heavy infection with the die-back fungus Calonectria rigidiuscula (Thresh, 1960). A few avirulent strains occur in limited widely scattered outbreaks, usually induce stem swellings only, and have little effect, if any, on growth or yield. Some virulent strains are widespread and lethal; in West African Amelonado cacao the severe New Juaben strain induces conspicuous swellings in stems and tap roots (Fig. 3, Fig. 4), necrosis of lateral roots (Fig. 4), defoliation, and the production of small rounded mottled pods (Fig. 6) containing fewer and smaller beans than normal pods, a transient red vein-banding on young leaves and interveinal chlorotic speckling and fern leaf patterns (Posnette, 1947) on mature leaves (Fig. 1, Fig. 2). Infection decreases yield by 25% within one year, 50%, within two years and usually kills trees within 3-4 years (Crowdy & Posnette, 1947). By contrast, Upper Amazon types of cacao imported into West Africa in 1944 from Trinidad and Brazil, particularly those originating from the Nanay (Posnette & Todd, 1951) and Iquitos (Dale, 1957) areas show resistance and tolerance to infection, intercrosses often producing highly tolerant progeny.
Geographical Distribution
Widespread in West Africa (Ghana, Ivory Coast, Nigeria, Togo Republic and Sierra Leone) and also occurs in Ceylon and possibly Sabah.
Host Range and Symptomatology
Experimental host range is limited to c. 30 species in the Bombacaceae, Tiliaceae, Sterculiaceae and Malvaceae, some showing differential susceptibility to different strains (Posnette, Robertson & Todd, 1950; Tinsley & Wharton, 1958). Natural host range is restricted to cacao, Cola chlamydantha, Ceiba pentandra, Cola gigantea var. glabrescens (Posnette et al., 1950), and Sterculia tragacantha (Legg & Agbodjan, 1969).
-
Diagnostic species
- Theobroma cacao
(cacao). Amelonado cacao is very susceptible (Posnette, 1947), cacao beans being readily infected by viruliferous mealybugs (Posnette & Strickland, 1948) and by mechanical inoculation with concentrated virus preparations (Brunt & Kenten, 1962); seedlings usually produce acute red vein banding and chlorotic leaf symptoms within 20-30 days and, 2-12 weeks later, swellings on shoots and tap roots and interveinal leaf chlorosis characteristic of the particular strain (Thresh & Tinsley, 1959). - Corchorus spp. These are hypersensitive and quickly killed by most strains
(Tinsley & Wharton, 1958).
- Most other hosts remain symptomless; some strains, however, induce transient chlorotic vein-banding leaf symptoms in Ceiba pentandra, Cola chlamydantha, and Cola gigantea, and conspicuous, permanent leaf chlorosis and severe stunting in Adansonia digitata (Posnette et al., 1950).
-
Propagation species
- Cacao is a suitable plant for maintaining cultures; cacao, Adansonia digitata and Bombax brevicuspe are convenient sources of virus for purification.
-
Assay species
- Cacao: by recording the proportion of inoculated Amelonado cacao beans producing infected seedlings.
Strains
Many distinct variants can be recognized; at one time they were named alphabetically but now, as originally (Posnette, 1947), they are usually named from the locality of their origin. The best known is the severe New Juaben strain (strain A; Theobroma virus 1A) which is widespread in the eastern region of Ghana (see Main Diseases). The characteristic features of other well known strains occurring in Ghana (e.g. Bisa, Bosomuoso, Bosumtwe, Mampong, mild New Juaben and Nkawkaw), Nigeria (e.g. Balogun, Ilesha, Elepo and Offa Igbo), Ivory Coast (e.g. Kongodia and Sankadiokro) and Ceylon have been summarized by Thresh & Tinsley (1959); Attafuah, Blencowe & Brunt (1963) describe those of a strain from Sierra Leone.
Transmission by Vectors
Transmissible by at least 14 mealybug species (Hemiptera: Coccidae). Planococcoides njalensis is often the most important, although in Nigeria P. njalensis, Planococcus kenyae and Planococcus citri are almost equally abundant. Nymphs (1st, 2nd and 3rd instars) and female adults are equally efficient vectors but male adults are unable to transmit. A pre-acquisition starvation period slightly increases vector efficiency, perhaps because it induces the insects to settle more quickly. The minimum acquisition feeding period is 20 min and the optimum, for long thought to be c. 50 min (Posnette & Robertson, 1950), is probably 2-4 days (Roivainen, 1969). There is no detectable latent period; insects may transmit within 15 min, but maximum transmission occurs after 2-10 hr (Posnette & Robertson, 1950; Dale 1955). The virus persists in feeding insects for 3 hr or less (Posnette & Strickland, 1948), but starved adults and 1st instar nymphs can retain virus for 49 and 24 hr respectively (Lister, 1953). No transmission occurs through eggs.
Vector specificity: Ferrisia virgata has transmitted all isolates tested except that from Mampong which, like cacao mottle leaf virus, is transmitted specifically by Pseudococcus longispinus (Posnette, 1950).
Transmission through Seed
The New Juaben and Bisa strains are not seed-borne (Posnette, 1947).
Transmission by Dodder
Not transmitted by Cuscuta chinensis (Posnette et al., 1950).
Serology
Antisera with titres of 1/128 have been prepared against strains from New Juaben, Mampong and Anibil; the low titres are probably due to the relatively low virus content of the preparations used for immunization. Gel-diffusion serological tests distinguish specific reactions from those attributable to host protein contaminants (Kenten & Legg, unpublished).
Relationships
The three strains tested show some antigenic differences, but are related to each other and to cacao mottle leaf virus (Kenten & Legg, unpublished).
Stability in Sap
Crude saps are non-infective but extracts made with antioxidants sometimes infect up to 5% of inoculated cacao beans (Brunt & Kenten, 1962); using concentrated virus preparations, transmission rates from 60 to 90% are obtained. In partially purified preparations, the thermal inactivation point (10 min) is c. 55-60°C, dilution endpoint 10-3-10-4 and infectivity is retained without appreciable loss for 2-3 months at 2°C (Brunt, Kenten & Nixon, 1964; Kenten & Legg, 1965).
Purification
Difficulties due to mucilaginous and polyphenolic material are overcome by extracting cacao leaves in ten to twenty times their weight of 0.05 M phosphate buffer at pH 6.0-8.0 containing added protein (1-2% w/v egg albumin, bovine serum albumin or, preferably, hide powder) in addition to either 0.05 M thioglycollate or 0.01-0.005 M diethyldithiocarbamate (Brunt & Kenten, 1963). Virus can then be sedimented by one or more cycles of differential centrifugation, or by ultracentrifugation (75,000 g for 1 hr) following a preliminary concentration by precipitation with ammonium sulphate (Kenten & Legg, 1965).
Properties of Particles
Sedimentation coefficient (s20,w) at infinite dilution of Mampong strain: 218 S.
Particle Structure
Particles are bacilliform (Fig. 5) about 121-130 x 28nm, with no discernible substructure when mounted in neutral 2% potassium phosphotungstate (Brunt et al., 1964; Kenten & Legg, unpublished). Empty square-ended tubular particles present in some preparations are probably degraded bacilliform particles (Kenten & Legg, 1967).
Particle Composition
No information.
Relations with Cells and Tissues
Tissues in chlorotic areas of leaves remain undifferentiated, contain small flattened chloroplasts and have reduced intercellular spaces; secondary xylem and phloem are both increased in stem and root swellings, but no phloem necrosis occurs (Knight & Tinsley, 1958).
Notes
Cacao swollen shoot and cacao mottle leaf viruses are both transmitted by mealybugs (Posnette, 1947), have similar particles and properties (Brunt et al., 1964; Kenten & Legg, 1967), and are now known to be serologically distantly related (Kenten & Legg, personal communication). It is uncertain, therefore, whether cacao mottle leaf virus should be considered a distinct virus or a serotype of cacao swollen shoot virus.
Two distinct virus strains transmissible by mealybugs infect cacao in Trinidad and another mealybug-transmitted virus infects pineapple in Hawaii (Carter, 1963); little is known about these and the viruses reported to infect cacao in Java, Colombia, Dominican Republic and Venezuela (Thresh & Tinsley, 1959) so that comparisons with cacao swollen shoot and mottle leaf viruses are not possible.
Figures

West African Amelonado cacao leaf with interveinal chlorosis induced by the severe New Juaben strain. (Photo courtesy A. F. Posnette.)

Chlorotic vein banding (‘fern leaf’ patterns) on mature Amelonado leaf infected with the New Juaben strain. (Photo courtesy A. F. Posnette.)

Stem swelling on naturally infected Amelonado tree. (Photo courtesy R. H. Kenten.)
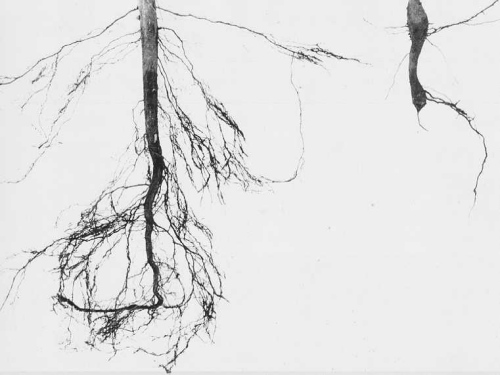
Root systems from 6-week-old seedlings; (left) healthy, (right) infected with the severe New Juaben strain.
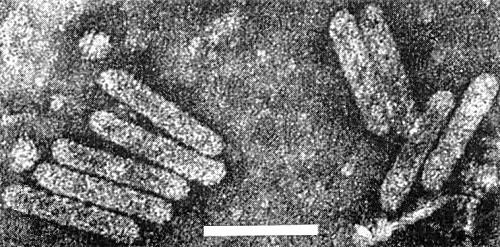
Virus particles from a partially purified preparation of the strain from Anibil, Ghana; mounted in 2% neutral potassium phosphotungstate. Bar represents 100 nm. (Photo courtesy R. D. Woods.)

(Left) small, rounded pod from severely affected tree, (right) normal pod. (Photo courtesy A. F. Posnette.)
References list for DPV: Cacao swollen shoot virus (10)
- Attafuah, Blencowe & Brunt, Trop. Agric. Trin. 40: 229, 1963.
- Brunt & Kenten, Ann. appl. Biol. 50: 749, 1962.
- Brunt & Kenten, Virology 19: 388, 1963.
- Brunt, Kenten & Nixon, J. gen. Microbiol. 36: 303, 1964.
- Carter, Ann. N.Y. Acad. Sci. 105: 741, 1963.
- Crowdy & Posnette, Ann. appl. Biol. 34: 403, 1947.
- Dale, Rep. W. Afr. Cocoa Res. Inst. 1954-55: 33, 1955.
- Dale, Rep. W. Afr. Cocoa Res. Inst. 1955-56: 35, 1957.
- Kenten & Legg, Ghana J. Sci. 5: 221, 1965.
- Kenten & Legg, J. gen. Virol. 1: 467, 1967.
- Knight & Tinsley, Ann. appl. Biol. 46: 7, 1958.
- Legg & Agbodjan, Rep. Cocoa Res. Inst. Ghana 1967-68: 25, 1969.
- Lister, Rep. W. Afr. Cocoa Res. Inst. 1952-53: 9, 1953.
- Posnette, Ann. appl. Biol. 34: 388, 1947.
- Posnette, Ann. appl. Biol. 37: 378, 1950.
- Posnette & Robertson, Ann. appl. Biol. 37: 363, 1950.
- Posnette & Strickland, Ann. appl. Biol. 35: 53, 1948.
- Posnette & Todd, Ann. appl. Biol. 38: 785, 1951.
- Posnette, Robertson & Todd, Ann. appl. Biol. 37: 229, 1950.
- Roivainen, Rep. Cocoa Res. Inst. Ghana 1967-68: 40, 1969.
- Thresh, Emp. J. exp. Agric. 28: 193, 1960.
- Thresh & Tinsley, Tech. Bull. W. Aft. Cocoa Res. Inst. 7: 32 pp., 1959.
- Tinsley & Wharton, Ann. appl. Biol. 46: 1, 1958.