Details of DPV and References
DPV NO: 100 October 1972
Family:
Genus:
Species: | Acronym:
Rice transitory yellowing
E. Shikata Department of Botany, Faculty of Agriculture, Hokkaido University, Sapporo, Japan
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
Described by Chiu et al. (1965).
A virus with bullet-shaped particles about 96 x 129 nm in dip preparations and 94 x 193 nm in ultrathin sections of diseased leaves. Rice is the only known host plant. The virus is transmitted in the persistent manner by rice green leafhoppers but not by inoculation of sap. Found only in Taiwan.
Main Diseases
Causes severe yellowing and stunting on the second rice crop in central and southern Taiwan.
Geographical Distribution
Taiwan.
Host Range and Symptomatology
Rice is the only known host. Echinochloa crusgalli and E. colonum plants are hosts for the insect vectors but are immune to the virus (Chiu et al., 1968). Rice varieties differ in symptom expression. Varieties of the japonica type are more susceptible and show more severe symptoms than do those of the indica type (Chiu et al., 1965; Su, 1969). A typical diseased plant of the variety Taichung 65, a japonica type, develops yellow or orange-buff colouring on the lower two or three leaves, yellowing on the distal half of the middle leaves and no symptom on the young leaves, 30 to 40 days after inoculation (Fig. 1, Fig. 2). Yellowing of the leaves starts from the tip of a leaf blade. Rusty flecks sometimes appear on the lower diseased leaves in the advanced stage of infection. Infection decreases the number of tillers (Fig. 1). In the greenhouse, diseased plants gradually recover and produce symptomless leaves (‘transitory yellowing’) (Fig. 2).
The virus can be assayed by injecting preparations into virus-free vector insects (Nephotettix cincticeps) and then allowing them to feed on Echinochloa colonum plants for 10 days before transfer to healthy rice (Taichung 65) test plants (Hsieh, 1967; Hsieh & Roan, 1967).
Strains
None described.
Transmission by Vectors
Transmitted by three species of rice green leafhoppers, Nephotettix apicalis, N. cincticeps (Chiu et al., 1968) (Fig. 3) and N. impicticeps (R. J. Chiu, pers. comm.). Insects can acquire the virus by feeding on diseased plants for 5 min to 10 h (N. apicalis) or 15 min to 4 h (N. cincticeps). The virus persists in its insect vectors for life after an incubation period from 3 to 29 days (N. apicalis) or 21 to 30 days (N. cincticeps). The virus does not pass through the leafhopper egg (Chiu & Jean, 1969).
Transmission through Seed
No information.
Serology
Antiserum with a moderately high titre was made by intravenous injection with partially purified virus, obtained according to Su (1969). In tube precipitin tests, the antiserum reacted with partially purified antigen from diseased plants, but not with antigen from healthy plants.
Relationships
The bullet-shaped particles of rice transitory yellowing virus distinguish it from other rice viruses (dwarf, stripe, black-streaked dwarf, tungro and hoja blanca). Morphologically it belongs to the rhabdovirus group (sensu Howatson, 1970), which contains at least 12 plant-infecting viruses. However, the particles of this virus are characteristic in being bullet-shaped in ultrathin sections as well as in dip preparations. Virus assembly is associated with nuclear membranes, in which respect it resembles potato yellow dwarf and sowthistle yellow vein viruses.
Stability in Sap
Properties of the virus were determined by injecting sap from diseased rice leaves into vector insects (N. cincticeps). The virus retained infectivity for 11 days at 0-2°C, and for 36 hr at room temperature (28-33°C). The thermal inactivation point of the virus was between 55.5°C and 57.5°C, and the dilution end-point between 10-5 and 10-6 (Hsieh, 1967).
Purification
The virus is sedimented by centrifugation at 10,000 rpm (6600 g) for 60 min (Hsieh & Roan, 1967), but purified preparations have not been obtained.
Properties of Particles
No information.
Particle Structure
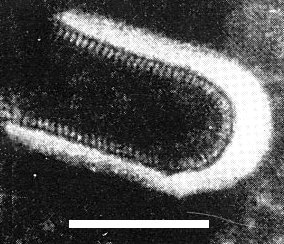
Bullet-shaped particles, 96 x 129 (range 120-140) nm are observed in negatively stained preparations of crude leaf extracts. Each particle has a deeply staining central core 45 nm wide; fine projections about 7 nm long extend outward from the outer membranes (Fig. 4). Cross striations 3 nm wide, about 5 nm apart, are shown (Fig. 5). In ultrathin sections of infected rice leaves, bullet-shaped particles 94 x 193 (range 180-210) nm were found (Shikata & Chen, 1969; Chen & Shikata, 1971) (Fig. 6).
Particle Composition
No information.
Relations with Cells and Tissues
Spherical or cylindrical inclusion bodies of various sizes appear mostly in the parenchyma cells around vascular bundles, also in the smaller parenchyma cells in the leaf and root. The chloroplasts in mesophyll cells disintegrate and have a decreased affinity for hematoxylin and Giemsa stains (Su, 1969). In ultrathin sections of diseased rice leaves, virus particles appear in the parenchyma cells bordering the vascular bundles and in the sieve elements. They occur in large numbers between the inner and outer lamellae of the nuclear membrane with their blunt ends attached to the inner nuclear membrane (Chen & Shikata, 1971). In viruliferous insects, bullet-shaped virus particles occur in the salivary gland, associated with the cytoplasmic membranes of the cells (Chen & Shikata, 1972).
Notes
The symptoms of transitory yellowing are indistinguishable from those of tungro of rice. However, the vector relations and the morphology of the two viruses differ. Tungro virus (Gálvez, 1971) is semi-persistent in its insect vector, Nephotettix impicticeps, and has spherical particles 30 nm in diameter.
Figures

Rice variety Taichung 65: (left) healthy, (right) infected with transitory yellowing virus, showing reduced tillering and discoloration of the leaves.

Infected leaves showing different degrees of discoloration. From right to left, orange-buff on a lower leaf, yellowing on a middle leaf, slight discoloration on the distal half of a middle leaf and no yellowing of a young leaf.

Insect vectors of rice transitory yellowing virus: (left) Nephotettix cincticeps, (right) N. apicalis; (above) female, (below) male.
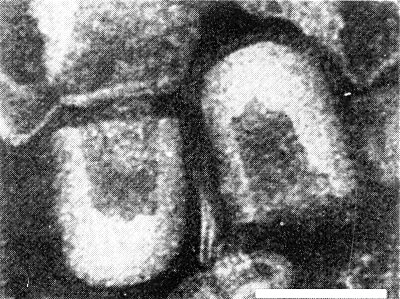
Virus particles from rice sap in 1% phosphotungstate. Bar represents 100 nm.
Virus particles from rice sap in 1% phosphotungstate. Bar represents 100 nm.
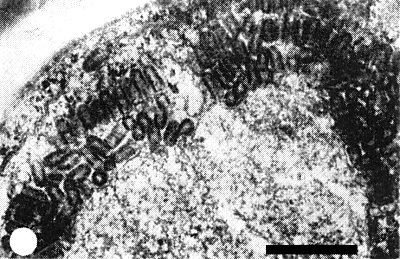
Virus particles aligned at the periphery of a nucleus of an infected rice cell. Bar represents 500 nm.
References list for DPV: Rice transitory yellowing (100)
- Chen & Shikata, Virology 46: 786, 1971.
- Chen & Shikata, Virology 47: 483, 1972:.
- Chiu & Jean, in The Virus Diseases of the Rice Plant, Compiled by Int. Rice Res. Inst., Johns Hopkins, Baltimore, p. 131, 1969.
- Chiu, Jean, Chen & Lo, Phytopathology 58: 740, 1968.
- Chiu, Lo, Pi & Chen. Bot. Bull. Acad. sin., Taipei 6: 1, 1965.
- Gálvez, CMI/AAB Descriptions of Plant Viruses 67: 3 pp., 1971.
- Howatson, Adv. Virus Res. 16: 196, 1970.
- Hsieh, Pl. Prot. Bull., Taiwan 9 (3-4): 21, 1967.
- Hsieh & Roan, Pl. Prot. Bull., Taiwan 9 (1-2): 23, 1967.
- Shikata & Chen, J. Virol. 3: 261, 1969.
- Su, in The Virus Diseases of the Rice Plant, compiled by Int. Rice Res. Inst., Johns Hopkins, Baltimore, p. 13, 1969.