Details of DPV and References
DPV NO: 111 July 1973
Family: Alphaflexiviridae
Genus: Potexvirus
Species: Clover yellow mosaic virus | Acronym: ClYMV
Clover yellow mosaic virus
L. Bos Institute of Phytopathological Research, Wageningen, The Netherlands
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
- Described by
Johnson (1942)
and
Pratt (1961).
Selected synonyms
- Marmor efficiens (Rev. appl. Mycol.
21: 293) - Pea mottle virus (Rev. appl. Mycol. 21: 293)
-
A virus with flexuous elongated particles c. 540 nm long. Readily transmissible by inoculation of sap to several legumes and some non-legumes, but no vector is known. Widely distributed in western North America.
Main Diseases
Causes yellow or light-green leaf striping (Fig. 1) in clovers, sometimes with stunting and leaf distortion; reduces winter hardiness and yield. Often occurs in complexes with the related white clover mosaic virus and with other viruses. Also found in lucerne, sweet clover, Chenopodium album and Stellaria media (Pratt, 1961; 1967), and in pea with necrotic streaking (Ford & Baggett, 1965). Welsh, Stace-Smith & Brennan, 1973 obtained clover yellow mosaic virus from apple trees with leaf pucker disease.
Geographical Distribution
Western USA and Canada (Pratt, 1961; Agrawal, Bos & Chessin, 1962).
Host Range and Symptomatology
Many species in the Papilionaceae are susceptible to systemic infection. Most isolates also systemically infect Antirrhinum majus, Chenopodium amaranticolor, Cucumis sativus, Gomphrena globosa, Spinacia oleracea, Stellaria media and Tetragonia expansa (Johnson, 1942; Pratt, 1961).
Diagnostic species
- Antirrhinum majus
(snapdragon). Local necrotic rings sometimes occur after 1-2 weeks; systemic chlorotic mottling and mild distortion, sometimes latent. (Not susceptible to white clover mosaic virus).- Chenopodium amaranticolor. Small chlorotic local lesions with necrotic centres
develop after 4-6 days, followed after 1-2 weeks by systemic fine mottling. (Not susceptible
to white clover mosaic virus).
- Cucumis sativus (cucumber). Transient chlorotic spots visible 1-4 weeks after inoculation. Many isolates infect systemically, as do some isolates of white clover mosaic virus.
- Gomphrena globosa. Distinct red rings with papery necrotic centres after 4-6 days (Fig. 2). With some isolates, severe systemic necrotic flecking. (Not susceptible to white clover mosaic virus).
- Pisum sativum (pea). Systemic vein chlorosis and chlorotic to yellow spotting after 6-12 days followed by slight stunting of the plant and retarded opening of new leaves (Fig. 3). Sometimes causes ‘streak’-like symptoms; no resistant varieties found (Ford & Baggett, 1965).
- Vicia faba (broad bean). Severe systemic mottling (Fig. 4) and sometimes mild necrotic flecking after 5-10 days. Occasionally small necrotic local lesions. One isolate studied caused symptomless infection.
- Vigna sinensis (cowpea). Symptoms indistinct; inoculated leaves become chlorotic and fall off; sometimes they develop small local lesions. In contrast, white clover mosaic virus induces local necrotic lesions or chlorotic spots or rings. Some clover yellow mosaic virus isolates infect systemically but more slowly than white clover mosaic virus.
- Cucumis sativus (cucumber). Transient chlorotic spots visible 1-4 weeks after inoculation. Many isolates infect systemically, as do some isolates of white clover mosaic virus.
Propagation species
- Pisum sativum
is a good source of virus for purification. To avoid possible contamination with white clover mosaic virus, systemic passage via Antirrhinum majus or C. amaranticolor is advisable.Assay species
- Chenopodium amaranticolor, Gomphrena globosa
and Phaseolus aureus give local lesions.
Strains
No special strains described, but 4 isolates differed slightly in symptom expression and serological affinity (Pratt, 1961).
Transmission by Vectors
The aphids Acyrthosiphon pisum and Anuraphis bakeri failed to transmit (Johnson, 1942; Pratt, 1961).
Transmission through Seed
Recorded in 5 out of 66 seedlings of Trifolium pratense (Hampton, 1963).
Transmission by Dodder
Cuscuta campestris (Johnson, 1942) was used to separate the virus (‘pea mottle’) from a complex with white clover mosaic virus (‘pea wilt’). Certain isolates are not readily transmissible (Pratt, 1961).
Serology
Antiserum titres of up to 1/2560 are readily obtained (Pratt, 1961; Bercks & Brandes, 1963). By concentrating the antibodies by precipitation with 40% ammonium sulphate, preparations with titres of 1/8192 and 1/32,768 were obtained (Bercks, 1963). With purified virus suspensions or clarified plant sap, precipitin tests in liquid media yield flocculent precipitates. In agar-gel diffusion tests, lines may develop after a long time (Ford, 1964a), but they are readily produced when virus fragmented with e.g. 0.1 M ethanolamine. pH 10.5, is used as antigen (Purcifull & Shepherd, 1964).
Relationships
Only minor serological differences were observed between different isolates of the virus (Pratt, 1961). It is distantly related serologically to cactus X, hydrangea ringspot, potato X, and white clover mosaic viruses, although all these viruses have slightly shorter particles (Bercks, 1963; Bercks & Brandes, 1963). All belong to the potexvirus group.
Stability in Sap
The thermal inactivation point is between 58 and 62°C, the dilution end-point around 10-4, and longevity in vitro exceeds 6 months, although infectivity considerably decreases between 6 and 12 months. One isolate was still infective after 22 months (Pratt, 1961).
Purification
Pisum sativum is most commonly used; purification from pea roots made it easier to obtain virus preparations free from chlorophyll, dark pigment and plant components (Ford, 1964b). The virus can be purified in various ways:
1. Homogenize fresh or frozen plant material in water or in 0.1 M phosphate buffer (pH 7.0), filter through cheesecloth and clarify by low speed centrifugation. After several cycles of differential centrifugation, resuspend pellets in buffer (Pratt, 1961; Agrawal, Chessin & Bos, 1962).
2. Grind fresh plant material in water, filter through cheesecloth, add 10% acetic acid to pH 4.5; after a few min add 10% ammonia to pH 8.0 and centrifuge at low speed. Decant the supernatant fluid through glass wool, adjust pH to 5.3 and remove precipitate by low speed centrifugation, resuspend pellet in 0.05 M borate pH 8.3 and clarify by low speed centrifugation; then dialyse for 24-48 h against 0.03 M neutral citrate in 0.01 M Tris-HCl buffer at pH 7.5. After 2 cycles of differential centrifugation, resuspend in water (Purcifull & Shepherd, 1964).
3. Clarify sap according to the ether-carbon tetrachloride method of Wetter (1960) and sediment and clarify by two cycles of differential centrifugation, resuspending the pellets in 0.01 M phosphate buffer (Bercks & Brandes, 1963).
Properties of Particles
The virus sediments as a single component in the analytical ultracentrifuge. Sedimentation coefficient at infinite dilution (in 0.1 M phosphate at pH 7.0) (s20): 121 S (for white clover mosaic virus: 112 S) (Pratt & Reichmann, 1961).
Isoelectric point about pH 5.3 (according to Purcifull & Shepherd (1964) maximum precipitation occurs at pH 5.3).
Electrophoretic mobility: -2.8 x 10-5 cm2 sec-1 volt-1 in 0.1 M phosphate buffer pH 7.5 (Purcifull & Shepherd, 1964).
A260/A280: 1.14 (calculated from curve given by Purcifull & Shepherd, 1964).
Absorbance at 260 nm (1 mg/ml, 1 cm light path): 3.1 (Purcifull & Shepherd, 1964).
Particle Structure
Particles are flexuous filaments c. 539 nm long (Fig. 5) (Bercks & Brandes, 1963). (Particles of white clover mosaic virus are c. 480 nm long).
Particle Composition
No information.
Relations with Cells and Tissues
In the first 400 µm of the root tip of Vicia faba no virus is detectable by local lesion assay on Gomphrena globosa (Smith & Schlegel, 1964). In many mesophyll cells of the chlorotic parts of red clover leaves, large inclusion bodies stainable with haematoxylin are visible by light microscopy (Pratt, 1966). Electron microscopy of ultrathin sections of systemically infected pea leaves reveals abundant inclusions of variable size and shape, consisting of aggregates of virus particles sectioned in various planes; some aggregates have a spiral structure (Fig. 6, Fig. 7) (Purcifull, Edwardson & Christie, 1966). By autoradiography of infected Vicia faba leaf tissue which had been treated with virus-specific labelled antibodies, structures containing virus protein were found in the cytoplasm and vacuoles; virus inclusions were only labelled at the edge (Schlegel & Delisle, 1971).
Notes
In western North America the virus often occurs together with white clover mosaic virus, from which it can be easily separated by systemic passage in Antirrhinum majus or Chenopodium amaranticolor. White clover mosaic virus (cf. Bercks, 1971) can be obtained free from clover yellow mosaic virus by early recovery from tip leaves of Vigna sinensis. The two viruses are distantly related serologically but differ in particle length. Other viruses often infecting clovers, such as alfalfa mosaic, bean yellow mosaic, pea streak and red clover vein mosaic, can be easily differentiated by host range, symptomatology, particle length and morphology.
Figures

Systemic symptoms in white clover. (After Pratt, 1961.)
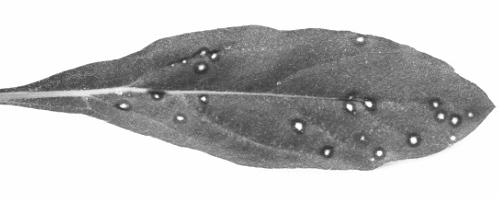
Local lesions in Gomphrena globosa. (After Pratt, 1961.)

Leaves of Pisum sativum, ‘Little Marvel’: (left) healthy; (right) infected, showing systemic symptoms. (After Pratt, 1961.)

Vicia faba leaves: (left) healthy; (right) infected, showing systemic mottle. (After Pratt, 1961.)

Electron micrograph of dip preparation; bar represents 500 nm. (After Brandes, 1964.)

Ultrathin section of virus aggregates in infected pea leaf tissue; bar represents 500 nm. (After Purcifull et al., 1966.)
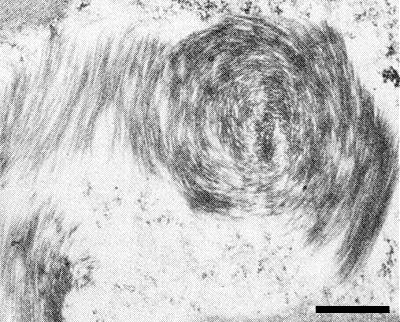
Ultrathin section of virus aggregates in infected pea leaf tissue; bar represents 500 nm. (After Purcifull et al., 1966.)
References list for DPV: Clover yellow mosaic virus (111)
- Agrawal, Bos & Chessin, Phytopathology 52: 517, 1962.
- Agrawal, Chessin & Bos, Nature, Lond. 194: 408, 1962.
- Bercks, Phytopath. Z. 47: 301, 1963.
- Bercks, CMI/AAB Descriptions of Plant Viruses 41: 4 pp., 1971.
- Bercks & Brandes, Phytopath. Z. 47: 381, 1963.
- Brandes, Mitt. biol. BundAnst. Ld- u. Forstw. 110, 130 pp., 1964.
- Ford, Phytopathology 54: 615, 1964a.
- Ford, Phytopathology, 54: 1433, 1964b.
- Ford & Baggett, Pl. Dis. Reptr 49: 787, 1965.
- Hampton, Phytopathology 53: 1139, 1963.
- Johnson, Phytopathology 32: 103, 1942.
- Pratt, Can. J. Bot. 39: 655, 1961.
- Pratt, Phytopathology 56: 895, 1966.
- Pratt, Can. J. Pl. Sci. 47: 289, 1967.
- Pratt & Reichmann, Proc. Can. phytopath. Soc. 28: 13, 1961.
- Purcifull, Edwardson & Christie, Virology 29: 276, 1966.
- Purcifull & Shepherd, Phytopathology 54: 1102, 1964.
- Schlegel & Delisle, Virology 45: 747, 1971.
- Smith & Schlegel, Phytopathology 54: 1273, 1964.
- Welsh, Stace-Smith & Brennan, Phytopathology 53: 50, 1973.
- Wetter, Arch. Mikrobiol. 37: 278, 1960.