Details of DPV and References
DPV NO: 115 July 1973
Family: Rhabdoviridae
Genus: Nucleorhabdovirus
Species: Eggplant mottled dwarf virus | Acronym: EMDV
Eggplant mottled dwarf virus
G. P. Martelli Istituto di Patologia Vegetale, Università di Bari, 70126 Bari, Italy
M. Russo Istituto di Patologia Vegetale, Università di Bari, 70126 Bari, Italy
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
Described by Martelli (1969) and Martelli & Cirulli (1969).
An enveloped virus with bacilliform particles c. 220 x 66 nm in plant tissue sections or c. 225 x 90 nm in partially purified preparations. It causes a severe disease of eggplant in Italy. Transmitted by grafting and by inoculation of sap.
Main Diseases
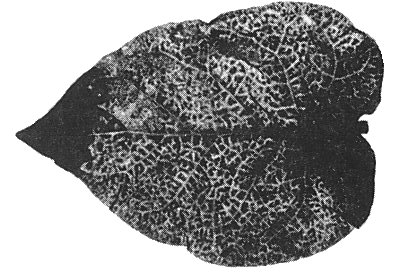
Causes severe stunting of eggplant (Solanum melongena) accompanied by pronounced mottling and crinkling of the leaves (Fig. 1) and general unfruitfulness. The low incidence in the field (only 1-2% of the plants are infected) limits the economic importance of the disease.
Geographical Distribution
Reported from southern Italy.
Host Range and Symptomatology
Eggplant is the only known natural host. Many varieties can be infected by grafting (Martelli & Cirulli, 1969) or mechanical inoculation (Martelli & Rana, 1970). Experimental host range is rather narrow, comprising species from four dicotyledonous families (Martelli & Rana, 1970). Readily transmitted by inoculation of sap to some species of Solanaceae (eggplant, Nicotiana species). Attempts to infect lettuce were unsuccessful.
-
Diagnostic species
- Solanum melongena
(eggplant). Small chlorotic local lesions after 2-3 weeks followed after another week by systemic mottle, severe vein-clearing, leaf deformation and crinkling (Fig. 1). - Nicotiana glutinosa. Chlorotic local lesions appear in inoculated leaves after
10-15 days. Systemic invasion is characterised by vein-yellowing (Fig. 2), flecking of
the leaves and stunting. Basal leaves wither and dry out.
- N. tabacum. Cvs. Samsun, White Burley and Xanthi-nc react like N. glutinosa but exhibit more obvious chlorotic local lesions (Fig. 3).
-
Propagation species
- S. melongena, N. glutinosa
and N. tabacum are suitable for maintaining cultures, and are good sources of virus for purification.Assay species
- N. tabacum,
and to a lesser extent N. glutinosa, can be used for local lesion assays.
Strains
No information.
Transmission by Vectors
The vector (probably aerial) is not known. Experimental transmission tests with aphids (Myzus persicae, Macrosiphum euphorbiae) and leafhoppers (Laodelphax striatellus) were unsuccessful (M. Conti, pers. comm.).
Transmission through Seed
No seed-transmission was found in N. glutinosa.
Transmission by Dodder
Not tested.
Serology
No information.
Relationships
In particle shape eggplant mottled dwarf virus resembles several other plant-infecting rhabdoviruses (sensu Howatson, 1970) but differs from all of them in one or more of the following characters: mode and site of particle development, particle size, transmissibility by inoculation of sap, symptom type and host range. It resembles lettuce necrotic yellows virus in the symptoms produced in some experimental hosts but differs in the mode of multiplication of the particles and in their intracellular localization (Wolanski & Chambers, 1971).
Stability in Sap
In sap of N. glutinosa or N. tabacum the thermal inactivation point (10 min) is about 54°C, dilution end-point is between 10-3 and 10-4 and infectivity is retained for 30-44 hr at 4°C (Martelli & Rana, 1970). The virus does not withstand desiccation or freezing in infected leaves. It is sensitive to organic solvents (n-butanol, chloroform, ether).
Purification
Clean virus preparations are obtained by grinding infected tissues in 0.2 M Na2HPO4 (pH 8.5) followed by filtration of expressed sap through Celite and differential centrifugation. However, the infectivity is lost during the early stages of purification (Russo & Martelli, 1973).
Properties of Particles
No information.
Particle Structure
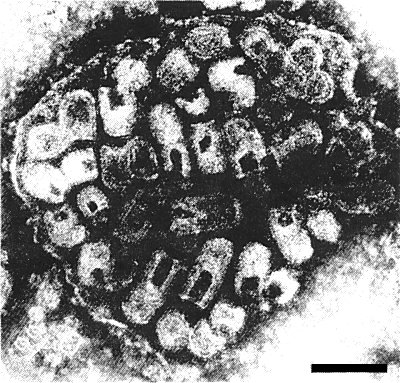
Bacilliform particles 220-230 x 90 nm are seen in partially purified preparations fixed in glutaraldehyde before negative staining with phosphotungstate. In unfixed samples and leaf dips mounted in phosphotungstate the particles are mostly bullet-shaped (Fig. 7) and measure 60-190 x 90 nm. Unfixed particles mounted in uranyl formate are mostly bacilliform. The virus particles exhibit surface projections c. 6 nm long and a helically arranged internal filament with a pitch of c. 4.5 nm (Fig. 6) which is thought to be the nucleocapsid. In cross section (Fig. 5) the particles show an outer envelope 10-12 nm thick which, depending on the fixing and staining procedure, may appear single- or double-layered. An electron-lucent region 5 nm across separates this external structure from an underlying ring c. 5.5 nm thick. The latter is, in turn, separated by an electron-lucent zone 4-4.5 nm wide from a central core, c. 18 nm in diameter, which itself has an electron-clear centre (Russo & Martelli, 1973). In tissue sections, ‘immature’ particles, i.e. those still connected to the nuclear envelope (Fig. 4), are bullet-shaped whereas the ‘mature’ particles are bacilliform and measure 220 x 66 nm (Martelli, 1969; Martelli & Castellano, 1970).
Particle Composition
No quantitative data available. Circumstantial evidence based upon the reaction of particles to differential electron microscope staining techniques suggests the presence of lipids in the outer envelope (Martelli & Castellano, 1970; Russo & Martelli, 1973). Pepsin-sensitive material constitutes the bulk of the particles (Russo & Martelli, 1973).
Relations with Cells and Tissues
The general structure of infected cells is fairly well preserved even when the plants show severe symptoms. The nuclei are the only organelles seriously affected, showing depletion of chromatinic material and uniformly granular nucleoplasm. Chloroplasts and mitochondria suffer minor structural modifications. Virus particles are present in leaf parenchyma tissue, in all parts of the flower except stamens, and in the fruit pericarp. They are also found in phloem parenchyma cells but not in mature sieve and tracheary elements. The particles develop by ‘budding’ at the nuclear envelope, with the nucleocapsid either oriented perpendicularly to the nucleus (Fig. 4) or, more rarely, parallel to the nuclear membrane. Virus particles accumulate in the perinuclear space. They can also be present in the ground cytoplasm within an extended network of membranous cisternae connected to the nuclear membrane (Martelli, 1969; Martelli & Castellano, 1970; Russo & Martelli, 1972).
Notes
Eggplant mottled dwarf virus is easily distinguished from other viruses infecting eggplant by the distinctive symptoms induced in the natural host and by the reaction of Nicotiana species to mechanical inoculation.
Figures

Systemic mottling and deformation in a leaf of naturally infected Solanum melongena.
Severe vein-yellowing in Nicotiana glutinosa.

Chlorotic local lesions in Nicotiana tabacum.

Cross section of the periphery of a nucleus (N) showing virus particles budding at the nuclear envelope into the spaces between the inner (il) and outer (ol) lamella of the membrane. Note the nuclear pore (np). Bar represents 200 nm.

A group of virus particles in cross section. Bar represents 150 nm.

Virus particles from a purified preparation mounted in phosphotungstate. Surface projections and transverse striations of the nucleocapsid are visible. Bar represents 200 nm.
A group of virus particles from a leaf dip preparation contained within an enveloping membrane. All the particles are bullet-shaped and the axial canal is partially penetrated by phosphotungstate. Bar represents 200 nm.
References list for DPV: Eggplant mottled dwarf virus (115)
- Howatson, Adv. Virus Res. 16: 195, 1970.
- Martelli, J. gen. Virol. 5: 319, 1969.
- Martelli & Castellano, Phytopath. medit. 9: 39, 1970.
- Martelli & Cirulli, Annls Phytopath. Numéro hors série: 393, 1969.
- Martelli & Rana, Phytopath. medit. 9: 187, 1970.
- Russo & Martelli, Phytopath. medit. 11: 136, 1972.
- Russo & Martelli, Virology 52: 39, 1973.
- Wolanski & Chambers, Virology 44: 582, 1971.