Details of DPV and References
DPV NO: 116 July 1973
Family: Potyviridae
Genus: Potyvirus
Species: Iris mild mosaic virus | Acronym: IMMV
There is a more recent description of this virus: DPV 324
Iris mild mosaic virus
A. A. Brunt Glasshouse Crops Research Institute, Littlehampton, Sussex, England
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
Disease described by
Van Slogteren (1963)
and
Loebenstein & Alper (1963);
virus partially
characterized by
Brunt (1968).
Selected synonyms
- Iris mosaic virus (Rev. appl. Mycol. 46: 350; 47: 397),
but now obsolete
(see Notes)
- Iris latent mosaic virus (Rev. appl. Mycol. 43: 162)
-
A virus with filamentous particles c. 760 x 12 nm. Transmissible by mechanical inoculation of sap, and in the non-persistent manner by aphids. Only known to occur naturally in bulbous irises, but transmissible to several other species. A typical, but apparently distinct member of the potyvirus group. World-wide distribution.
Main Diseases
Causes a mild mosaic disease of bulbous irises, and is especially prevalent in Iris xiphioides (English iris), I. xiphium (Spanish iris), I. tingitana x I. xiphium (Dutch iris), I. reticulata and I. danfordiae.
Geographical Distribution
World-wide; most cultivars that are commercially important are probably totally infected (Van Slogteren, 1963; Loebenstein & Alper, 1963; Lawson, 1967).
Host Range and Symptomatology
Iris spp. are the only known natural hosts, but the virus can also infect 9 species in 4 dicotyledonous families (Brunt, 1968, and unpublished).
-
Diagnostic species
- Iris
spp. Light green mosaic symptoms, often inconspicuous on leaves but slightly more intense on spathes (Fig. 1); flowers usually unaffected (Van Slogteren, 1963; Loebenstein & Alper, 1963; Lawson, 1967). - Chenopodium amaranticolor. A few circular necrotic lesions 2-4 mm in
diameter after 14-21
days
(Fig. 3);
no systemic infection.
- Nicotiana clevelandii. Inoculated leaves usually develop faint chlorotic lesions after 10-15 days, followed by faint systemic interveinal chlorosis (Fig. 4).
- Nicotiana megalosiphon. Inoculated leaves are usually symptomless; systemically infected leaves develop faint interveinal chlorosis after 14-28 days.
- Tetragonia expansa. Circular chlorotic local lesions after 11-14 days, slowly enlarging to 4-8 mm in diameter with dark green or grey necrotic margins (Fig. 2).
- Nicotiana clevelandii. Inoculated leaves usually develop faint chlorotic lesions after 10-15 days, followed by faint systemic interveinal chlorosis (Fig. 4).
-
Propagation species
- Iris
spp. For maintenance of cultures. - Nicotiana clevelandii and N. megalosiphon are good sources of inoculum and of virus for purification.
-
Assay species
- Tetragonia expansa,
although not highly sensitive, is a useful local lesion assay host.
Strains
None distinguished.
Transmission by Vectors
Transmissible in the non-persistent manner by Aphis gossypii and Myzus persicae, but not by Macrosiphoniella sanborni. All instars can transmit. Aphids acquire and inoculate virus in 5-10 min feeding periods; no latent period (Brunt, unpublished).
Transmission through Seed
Not seed-borne in iris (Brunt, 1968) or Nicotiana clevelandii.
Transmission by Dodder
No reports.
Serology
The virus is a moderately good immunogen; antisera, obtained from rabbits injected on three occasions with a total of c. 0.15 mg of virus, had homologous titres of 1/1024 in tube precipitin tests (Brunt, 1968), and antisera obtained after multiple intravenous injections had similar titres in microprecipitin tests (D. H. M. Van Slogteren, unpublished).
Relationships
In particle morphology and other properties, the virus resembles potato Y and allied viruses. However, it seems serologically unrelated to iris severe mosaic (Van Slogteren, 1963; Lawson, 1967), bearded iris mosaic (Barnett, De Zoeten & Gaard, 1971), potato Y or bean yellow mosaic viruses, nor to 14 other morphologically similar viruses (Brunt, 1968, and unpublished), and thus seems to be a typical but distinct member of the potato virus Y group.
Stability in Sap
In Tetragonia expansa sap, the thermal inactivation point (10 min) is 65-70°C, dilution end-point 10-3 to 10-4, and infectivity is retained for 3-4 days at 20°C and 16-32 days at 2°C. Sap lyophilized with dextrose and peptone remains infective for at least 2 years (Brunt, 1968).
Purification
The following procedure yields c. 5-10 mg of virus per kg of Nicotiana clevelandii leaf tissue. ‘Homogenize’ systemically infected leaves in 0.1 M borate buffer containing 0.05 M disodium ethylenediamine-tetraacetate and 0.2% 2-mercaptoethanol (pH 7.8-8.0), squeeze through cheesecloth and centrifuge the expressed fluid briefly at 10,000 g; shake the supernatant fluid with 50% (v/v) chloroform, sediment virus from the aqueous phase by centrifuging at 75,000 g for 90 min, and resuspend pellets in a small volume of 0.05 M borate buffer (pH 7.8). The virus is further purified by removing and concentrating the specific virus zones formed by centrifuging partially purified virus preparations in 10-40% (w/v) sucrose density gradient columns. Substantial amounts of virus can often be recovered from the plant fibre that is retained after squeezing the extract through cheesecloth by repeating the procedure.
Properties of Particles
Preparations usually contain one sedimenting component with a sedimentation coefficient
(s20, w) of 154 S, but the virus occasionally forms faster sedimenting
aggregates.
A260/A280: 1.25.
A260(max)/A247(min): 1.27 (after correction for light-scattering).
Particle Structure
Particles (Fig. 5) are slightly flexuous filaments c. 760 x 12 nm (Brunt, 1968). Uranyl formate penetrates particles to reveal a narrow central canal and some surface structure.
Particle Composition
RNA: c. 5-6% of particle weight (estimated spectrophotometrically).
Protein: c. 95% of particle weight.
Relations with Cells and Tissues
No amoeboid intracellular inclusions are present in infected iris or N. clevelandii cells. Sap, however, contains fragments of pinwheel plates (Brunt, unpublished).
Notes
Brierley & McWhorter (1936) first suspected that two viruses occurred in bulbous irises; later, Van Slogteren (1963) found that two serologically distinct viruses infected irises in the Netherlands, and Loebenstein & Alper (1963) reported the occurrence of two viruses in irises in Israel. Unfortunately, the name ‘iris mosaic virus’ has been used for both viruses: in North America it was used frequently for a virus causing severe leaf chlorosis, flower discoloration and stunting (see Martyn, 1968), whereas in western Europe it was used for the very prevalent but much less damaging virus described here (Van Slogteren, 1963; Baruch & Quak, 1966; Brunt, 1968). Further confusion can probably be best avoided by calling these viruses iris severe mosaic and iris mild mosaic, respectively (Lawson, 1967). Both viruses are aphid-borne and have filamentous particles, but they are serologically distinct and can be separated in differential hosts: iris mild mosaic virus, unlike iris severe mosaic virus, induces mild systemic chlorosis in Nicotiana clevelandii and Nicotiana megalosiphon, and local lesions without subsequent systemic infection in Tetragonia expansa, whereas only iris severe mosaic virus induces chlorotic local lesions in Chenopodium quinoa and the formation of amoeboid intracellular inclusions in irises.
Tobacco ringspot and tobacco rattle viruses also infect bulbous irises (Asjes, 1969), but are readily distinguished from iris severe mosaic and iris mild mosaic viruses by host range, symptomatology and other properties.
Iris mild mosaic virus is now so prevalent that virus-free stocks of commercially important cultivars are probably unobtainable by selection and roguing; although bulbs cannot be freed of virus by heat-treatment (Kassanis & Broadbent, 1958), virus-free plants obtained by meristem tip culture in Britain (Stone & Hollings, 1965) and in the Netherlands (Baruch & Quak, 1966) form foundation stocks of healthy plants.
Figures

Mild systemic chlorosis on leaves and spathe of naturally-infected ‘Wedgwood’ iris.
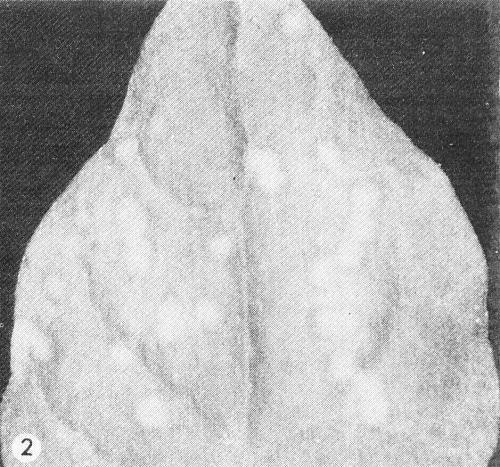
Inoculated Tetragonia expansa leaf with chlorotic lesions.
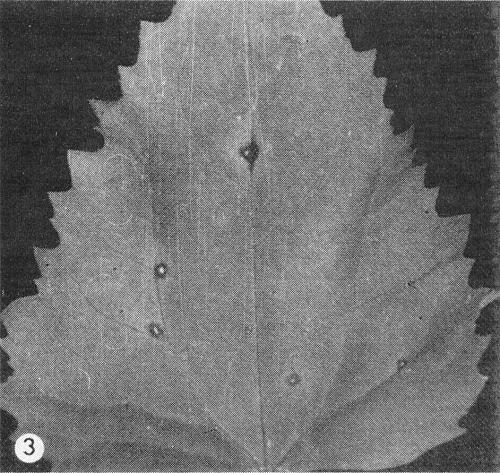
Inoculated Chenopodium amaranticolor leaf with few necrotic lesions.

Mild systemic leaf chlorosis in Nicotiana clevelandii.

Virus particles from a partially purified preparation mounted in neutral phosphotungstate. Bar represents 300 nm.
References list for DPV: Iris mild mosaic virus (116)
- Asjes, Versi. Werkzaamh. Bloembollen Onderz. Lisse 1968-69: 44, 1969.
- Barnett, De Zoeten & Gaard, Phytopathology 61: 926, 1971.
- Baruch & Quak, Neth. J. Pl. Path. 72: 270, 1966.
- Brierley & McWhorter, J. agric. Res. 53: 621, 1936.
- Brunt, Ann. appl. Biol. 61: 187, 1968.
- Kassanis & Broadbent, Rep. Rothamsted exp. Stn 1957: 106, 1958.
- Lawson, Pl. Dis. Reptr 51: 952, 1967.
- Loebenstein & Alper, Phytopathology 53: 349, 1963.
- Martyn, Commonw. Phytopath. Papers 9: 56, 1968.
- Van Slogteren, Versl. Werkzaamh. Bloembollen Onderz. Lisse 1962: 80, 1963.
- Stone & Hollings, Rep. Glasshouse Crops Res. Inst. 1964: 95, 1965.