Details of DPV and References
DPV NO: 119 July 1973
Family: Reoviridae
Genus: Fijivirus
Species: Fiji disease virus | Acronym: FDV
Sugarcane Fiji disease virus
P. B. Hutchinson C.S.R. Research Laboratories, Roseville, New South Wales, Australia
R. I. B. Francki Department of Plant Pathology, Waite Agricultural Research Institute, University of Adelaide, South Australia
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
First described by
Lyon (1910).
Synonyms
- Saccharum
virus 2 (Rev. appl. Mycol. 17: 52) - Galla fijiensis (Rev. appl. Mycol. 28: 512)
-
A virus with polyhedral particles c. 70 nm in diameter, probably containing double-stranded RNA. Its host range seems restricted to a few genera of the Gramineae. It is not transmissible by sap inoculation but is transmitted in a persistent manner by leafhoppers (Perkinsiella spp.).
Main Diseases
Causes ‘Fiji disease’ (Fig. 1, Fig. 2, Fig. 3) with severe stunting, deformation and death in susceptible varieties of sugarcane (species and hybrids within the genus Saccharum).
Geographical Distribution
Australia, Fiji, Madagascar, New Britain, New Guinea, New Hebrides, Philippines, Samoa.
Host Range and Symptomatology
All known hosts are members of the Gramineae. The virus is not transmissible by inoculation of sap (Hughes & Robinson, 1961) but can be transmitted by leafhopper vectors to sugarcane, other Saccharum spp., maize (Zea mays) and Sorghum spp. (Hutchinson, Forteath & Osborn, 1972).
Infected plants are stunted, with elongated swellings or galls along the veins on the lower surfaces of the leaves and leaf sheaths (Fig. 2). A conspicuous symptom in the field is the deformation and ‘bitten’ appearance of the leaves unfolding from the spindle (Fig. 3).
-
Diagnostic species
- Highly susceptible varieties of sugarcane, e.g. Fiji-10, H47-4991. Following exposure to viruliferous leafhoppers, the leaves of young plants develop galls after a minimum period of about 18 days. There is a strong correlation between the field resistance of a variety and the number of days between infection and the appearance of symptoms (Daniels et al., 1969).
Propagation species
- Virus can be maintained in sugarcane plants. Plants from infected setts develop symptoms on new leaves.
Assay species
- Susceptible varieties of sugarcane (e.g. Fiji-10) are used as systemic assay hosts.
Strains
No distinct strains have been recognized, but sugarcane varieties that are resistant in one country may not be resistant in others. This may reflect strain differences in the virus or, conceivably, the leafhopper vector.
Transmission by Vectors
Transmissible by the leafhoppers Perkinsiella saccharicida. P. vitiensis and P. vastatrix in a persistent manner. Present evidence suggests that the virus cannot be acquired after the first nymphal instar (P. B. Hutchinson, unpublished data). Once acquired, the virus can be transmitted throughout the life of the insect at any subsequent stage of the life cycle. It is not known whether the virus is transmitted through the egg. Infective insects for experimental work are obtained only from populations reared for a complete generation on infected plants. The virus probably multiplies in vector tissue (Francki & Grivell, 1972). An inoculation feeding time of 24 hr is used routinely in experimental work but it is likely that much shorter feeding times are adequate.
Transmission through Seed
No evidence suggesting transmission.
Transmission by Dodder
Not tested.
Serology
Antisera have not been prepared.
Relationships
Little is known about the relationships of the virus, but its structure resembles that of wound tumor, rice dwarf and maize rough dwarf viruses (Teakle & Steindl, 1969; Francki & Grivell, 1972).
Stability in Sap
Not investigated.
Purification
Not yet achieved.
Properties of Particles
Not investigated.
Particle Structure
Polyhedral particles about 70 nm in diameter occur in sections of infected cells of plants and vector insects (Fig. 5, Fig. 6) and in negatively stained leaf-dip preparations (Fig. 4) (Teakle & Steindl, 1969; Francki & Grivell, 1972).
Particle Composition
Double-stranded RNA was detected by a serological method (Francki & Jackson, 1972) in extracts of gall tissue from infected sugarcane but not in extracts from tissues of healthy plants. The double-stranded RNA was detected only in extracts that were deproteinized with phenol/sodium dodecyl sulphate, suggesting that it originated from virus particles (Francki & Jackson, 1972).
Relations with Cells and Tissues
Infected cells contain intracellular bodies visible by light microscopy (Kunkel, 1924). In the electron microscope, particles can be observed in the cytoplasm of gall cells in the vascular bundles of infected leaves (Fig. 5). The cells also contain inclusions resembling ‘viroplasm’ (Teakle & Steindl, 1969; Francki & Grivell, 1972). Similar particles and ‘viroplasm’ have been observed in cells of the leafhopper vector, P. saccharicida (Fig. 6). The distribution and appearance of particles and the presence of ‘viroplasm’ suggest that the virus multiplies in vector tissues (Francki & Grivell, 1972).
Notes
The leaf-galls are diagnostic and distinguish Fiji disease from other diseases of sugarcane except perhaps the leaf-galls recorded in Kenya (Sheffield, 1969) which are caused by an insect toxin. There are no leaf spot or stripe symptoms such as occur with sugarcane mosaic virus. The virus cannot be controlled by heat treatments which are effective against chlorotic streak or ratoon stunting disease. Some control can be achieved by the use of disease-free seed-beds, intensive roguing and insecticides but the only permanent control lies in the use of resistant varieties. Highly resistant varieties are available but are not commercially acceptable in some areas for other reasons. Many thousands of varieties have been tested for resistance to the disease and information on the results of these tests can be obtained through the INCANDEX system of the International Society of Sugarcane Technologists (Hutchinson, 1969).
Figures

Fiji disease in sugarcane. Complete loss of crop in a plot of a highly susceptible variety. Note the resistant variety in the background.

Sugarcane. Galls on the underside of a young infected leaf.

Sugarcane. Infected leaves with typical ‘bitten’ leaf symptoms.

Polyhedral particles in leaf-dip preparations of gall tissue from infected sugarcane leaves. Bar represents 100 nm.
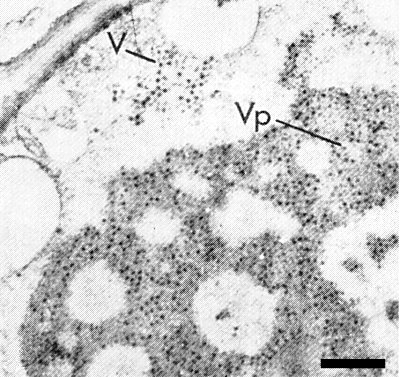
Typical inclusions in leaf gall cells. V = virus particles in the cytoplasm; Vp = virus particles embedded in ‘viroplasm’. Bar represents 1 µm.

Virus-infected fat-body cell of Perkinsiella saceharicida. Vp = virus particles embedded in ‘viroplasm’; Cr = crystalline arrangement of virus particles. Bar represents 1 µm.
References list for DPV: Sugarcane Fiji disease virus (119)
- Daniels, Ayub Husain, Hutchinson & Wismer, Proc. int. Soc. Sug. Cane Technol. 13: 1100, 1969.
- Francki & Grivell, Virology 48: 305, 1972.
- Francki & Jackson, Virology 48: 275, 1972.
- Hughes & Robinson, In Sugarcane diseases of the World, p. 389, Eds J. P. Martin, E. V. Abbott & C. G. Hughes, Elsevier, 1961.
- Hutchinson, Sugcane Path. Newsl. 3: 7, 1969.
- Hutchinson, Forteath & Osborn, Sugcane Path. Newsl. 9: 12, 1972.
- Kunkel, Bull. Hawaiian Sug. Plrs' Ass. Exp. Stn Bot. Ser. 3: 99, 1924.
- Lyon, Hawaii. Plrs’ Rec. 3: 200, 1910.
- Sheffield, Sugcane Path. Newsl. 3: 25, 1969.
- Teakle & Steindl, Virology 37: 139, 1969.