Details of DPV and References
DPV NO: 124 July 1973
Family: Tymoviridae
Genus: Tymovirus
Species: Eggplant mosaic virus | Acronym: EMV
Eggplant mosaic virus
A. J. Gibbs Research School of Biological Sciences, Australian National University, Canberra, Australia
B. D. Harrison Scottish Horticultural Research Institute, Invergowrie, Dundee, Scotland
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
Described by
Ferguson (1951) and
Dale (1954).
Synonym
- Andean potato latent virus (a distinctive strain) (Rev. appl. Mycol. 46: 78)
-
An RNA-containing virus with isometric particles of about 30 nm diameter, occurring in the West Indies and South America. It often occurs at high concentration in infected leaves, is readily sap-transmissible and has a beetle vector.
Main Diseases
The type strain causes mosaic of eggplant (Solanum melongena) (Fig. 1) and tomato (Lycopersicon esculentum). The Andean potato latent (APL) strain infects wild potato (Solanum tuberosum) symptomlessly.
Geographical Distribution
Trinidad, West Indies (type strain). Bolivia, Colombia and Peru (APL strain; Gibbs et al., 1966).
Host Range and Symptomatology
Host range includes about 30 species, mostly in the Solanaceae or Chenopodiaceae (Dale, 1954; Gibbs et al., 1966; Gibbs & Harrison, 1969). Readily transmissible by inoculation of sap.
-
Diagnostic species
- Chenopodium amaranticolor
and C. quinoa. Chlorotic or necrotic lesions in inoculated leaves. Systemic mottle or mosaic. - Cucumis sativus (cucumber). Inoculated leaves become infected symptomlessly. Not
systemic.
- Nicotiana clevelandii. Necrotic (type strain) or chlorotic (APL strain) lesions in inoculated leaves (Fig. 2). Systemically infected leaves develop mottling with or without necrosis. Plants infected with the type strain become yellowish and remain very stunted.
- N. glutinosa. Chlorotic or necrotic (Fig. 3) lesions develop in inoculated leaves. Systemically infected leaves develop chlorotic lesions and necrotic flecks, with or without distortion.
- Phaseolus vulgaris cv. The Prince (French bean). Not infected.
- Nicotiana clevelandii. Necrotic (type strain) or chlorotic (APL strain) lesions in inoculated leaves (Fig. 2). Systemically infected leaves develop mottling with or without necrosis. Plants infected with the type strain become yellowish and remain very stunted.
-
Propagation species
- Cultures can be maintained in Nicotiana clevelandii or N. glutinosa. N. clevelandii is a good source of virus for purification.
-
Assay species
- The type strain produces countable local lesions in Chenopodium amaranticolor. Dale (1954) reported that local lesions develop in C. hybridum and Vigna sinensis cv. Black-eye (cowpea).
Strains
Type strain (Dale, 1954). Accumulates best in leaves at 20-25°C.
Andean potato latent strain (Gibbs et al., 1966). This is less virulent than the type strain and accumulates best in leaves at 15°C. It differs serologically from the type strain (see below) and was found in wild potatoes collected from the tropical Andes.
Variants that differ in virulence but not serologically from the type strain are reported from Trinidad (Gibbs & Harrison, 1969).
Transmission by Vectors
Flea-beetles (Epitrix sp.) transmit the type strain, but no details of vector relations are available.
Transmission through Seed
Obtained in Nicotiana clevelandii and Petunia hybrida (APL strain). The infected seedlings looked normal (Gibbs et al., 1966).
Transmission by Dodder
No reports.
Serology
Strongly immunogenic in rabbits. Antisera with precipitin titres of 1/1000 are easily obtained. Precipitation tests in 1% agar gel are convenient for detecting the virus in crude sap; the virus gives one line of precipitate.
Relationships
Many of the antibodies (10 to 50%, depending on the antiserum) to the type and APL strains react with the heterologous strain, but the two are readily distinguished by spur formation in gel diffusion tests (Gibbs & Harrison, 1969; Bercks et al., 1971). Both strains are distantly serologically related to belladonna mottle, dulcamara mottle, ononis yellow mosaic and scrophularia mottle viruses (Bercks et al., 1971) which together make up one subgroup of the tymoviruses (Gibbs et al., 1966). Eggplant mosaic virus also shares many properties with other tymoviruses but no serological relationship was detected to turnip yellow mosaic virus itself.
In plant protection tests no protection occurred between the APL strain and either dulcamara mottle or ononis yellow mosaic viruses.
Stability in Sap
In Nicotiana glutinosa sap, the APL strain usually lost infectivity after 10 min at 90°C, storage at room temperature for a week, or dilution to 10-7.
Purification
Disrupt infected leaves in 0.1 M phosphate buffer, pH 7.0 (1 ml/2 g leaf), containing 0.05 M ascorbic acid, using a mechanical blender. Add chloroform and n-butanol (each one quarter of the volume of extract). Blend again to form an emulsion and then centrifuge at 8000 g for 10 min. Collect the aqueous phase and concentrate and purify the virus further by two or more cycles of differential centrifugation (100,000 g for 2 h and 8000 g for 10 min) using 0.03 M phosphate buffer, pH 7.0 to resuspend the virus-containing pellets obtained by high speed centrifugation (Gibbs et al., 1966).
Properties of Particles
The particles are all the same size but sediment as two components (Fig. 5), empty protein shells (T) and nucleoprotein particles (B).
Sedimentation coefficient (s20, w) at infinite dilution (svedbergs): 53 (T), 111 (B).
Particle Structure
Icosahedral symmetry, about 30 nm in diameter. Electron micrographs of negatively stained virus show 32 morphological subunits (Fig. 4) and the structure is probably the same as that of turnip yellow mosaic virus.
Particle Composition
RNA: Single-stranded, about 36% of the particle weight (calculated from the sedimentation coefficients of T and B particles). M. Wt, estimated from electrophoretic mobility in polyacrylamide gels, about 2.5 x 106 (Bercks et al., 1971), but estimated from particle structure and composition about 2.0 x 106.
Protein: About 64% of the particle weight. One species of protein in the particle; M. Wt 20,500 when estimated from electrophoretic mobility in polyacrylamide gels (Bercks et al., 1971; A. J. Gibbs & P. W. MacDonald, unpublished results), but 500 daltons less when estimated from the composition, of which the best integer fit is ala 25, arg 4, asp 16, cys 1, glu 13, gly 7, his 3, ile 13, leu 20, lys 7, met 3, phe 6, pro 19, ser 20, thr 20, try (not done), tyr 4, val 18, total 199 (A. J. Gibbs & P. W. MacDonald, unpublished results).
Relations with Cells and Tissues
In N. clevelandii leaves the chloroplasts develop marginal vesicles (Fig. 6) resembling those induced in Chinese cabbage by turnip yellow mosaic virus. This effect seems typical of tymoviruses. Occasionally a crystal composed of virus particles is found in the cell vacuole (B. D. Harrison & I. M. Roberts, unpublished results).
Notes
Eggplant, potato and tomato are subject to many different virus infections. Eggplant mosaic virus can be distinguished from many of these viruses by the size and shape of its particles. It also differs from most if not all in having the following combination of properties: sap-transmissible, systemically infecting Chenopodium amaranticolor, C. quinoa and Nicotiana glutinosa, infecting only the inoculated leaves of Cucumis sativus and failing to infect Phaseolus vulgaris. Additional characteristics useful for differentiation are its high thermal inactivation point and high concentration in tissues. Eggplant mosaic virus seems commoner in eggplant in Trinidad than cucumber mosaic or tobacco mosaic viruses. In the tropical Andes, the APL strain occurred in potato in mixed infections with potato S, potato X, potato Y and potato leaf roll viruses.
Acknowledgements
Figs 1-5 from Gibbs & Harrison, 1969; Fig. 6 courtesy I. M. Roberts.
Figures

Systemically infected leaf of Solanum melongena.

Inoculated leaves of Nicotiana clevelandii; (left) type strain, (right) APL strain.

Nicotiana glutinosa leaf inoculated with the type strain.

Virus particles treated with uranyl formate. Bar represents 100 nm.
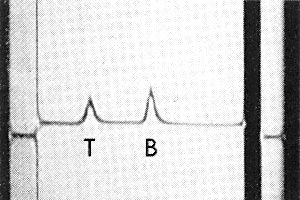
Schlieren diagram obtained by analytical ultracentrifugation of a purified virus preparation showing top (T) and bottom (B) components. Sedimentation from left to right.
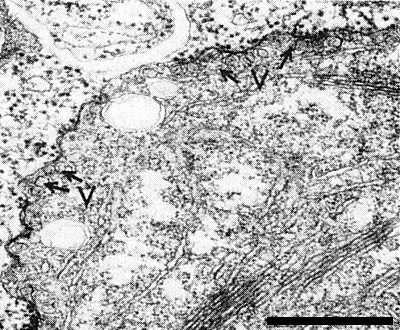
Section of N. clevelandii chloropast, showing marginal vesicles (V). Bar represents 500 nm.
References list for DPV: Eggplant mosaic virus (124)
- Bercks, Huth, Koenig, Lesemann, Paul & Querfurth, Phytopath. Z. 71: 341, 1971.
- Dale, Ann. appl. Biol. 41: 240, 1954.
- Ferguson, Pl. Dis. Reptr 35: 102, 1951.
- Gibbs & Harrison, Ann. appl. Biol. 64: 225, 1969.
- Gibbs, Hecht-Poinar, Woods & McKee, J. gen. Microbiol. 44: 177, 1966.