Details of DPV and References
DPV NO: 128 July 1974
Family: Tymoviridae
Genus: Tymovirus
Species: Okra mosaic virus | Acronym: OkMV
Okra mosaic virus
Louise Givord Laboratoire de Virologie, Centre ORSTOM d'Adiopodoumé, B.P. 20 - Abidjan, Côte d'Ivoire
Renate Koenig Biologisehe Bundesanstalt für Land-und Forstwirtschaft, Braunschweig, Germany
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
Described by
Givord, Pfeiffer & Hirth (1972) and
Givord & Hirth (1973).
Synonym
- Virus de la mosaïque du gombo (Rev. Pl. Path. 52: 551)
-
An RNA-containing virus with isometric particles c. 28 nm in diameter which sediment as two components in the ultracentrifuge. It has a wide host range and is readily transmitted by sap inoculation. No vector known. Found in West Africa.
Main Diseases
Causes a mosaic disease in okra (Hibiscus esculentus) (Fig. 1, Fig. 2, Fig. 3, Fig. 4) (Givord et al., 1972) and spotting in H. rosa-sinensis (L. Givord, unpublished).
Geographical Distribution
Recorded from Ivory Coast (Givord et al., 1972) and Nigeria (L. Givord, unpublished data). Probably widespread in West Africa.
Host Range and Symptomatology
Readily transmissible by inoculation of sap; it infected 105 species and varieties in 23 dicotyledonous families tested (Givord & Hirth, 1973; L. Givord, unpublished data).
-
Diagnostic species
- Hibiscus esculentus
(okra) cv. Clemson Spineless. Systemic symptoms appear on the youngest leaf about 7-8 days after inoculation, as a light green mosaic or regular veinal chlorosis (Fig. 1, Fig. 2). On the next two or three leaves produced, one or more of the principal veins are bordered by broad chlorotic bands (Fig. 3, Fig. 4). Leaves produced subsequently show no symptoms. - Arachis hypogaea (groundnut). Systemic dark and light green mosaic
(Fig. 6).
- Chenopodium amaranticolor. Systemic chlorotic spotting and systemic line pattern.
- Cucumis sativus (cucumber). Large chlorotic local lesions sometimes appear in inoculated cotyledons; fine vein chlorosis develops on the succeeding leaves.
- Nicotiana clevelandii. Systemic dark and light green and whitish mosaic, dwarfing.
- Vigna sinensis (cowpea). Systemic fine, regular vein chlorosis (Fig. 5) and mosaic.
- Chenopodium amaranticolor. Systemic chlorotic spotting and systemic line pattern.
-
Propagation species
- Hibiscus esculentus
cv. Clemson Spineless is a suitable plant for maintaining cultures in tropical areas. In temperate regions, cultures can be maintained in Cucumis sativus. Both species are good sources of virus for purification.Assay species
- No reliable local-lesion host known.
Strains
Ivory Coast isolate from okra (Givord & Hirth, 1973) systemically infects Vinca rosea, Cucumis sativus cv. Vert Long Maraîcher and Chenopodium amaranticolor, but is not transmissible to Phaseolus vulgaris cv. The Prince, Nicotiana tabacum cv. Samsun or N. glutinosa.
Ivory Coast isolate from Hibiscus rosa-sinensis (L. Givord, unpublished data) produces a mild mosaic in okra. It does not infect V. rosea, C. sativus cv. Vert Long Maraîcher, C. amaranticolor, P. vulgaris cv. The Prince or N. tabacum cv. Samsun but causes local infection in N. glutinosa.
Nigerian okra isolate (L. Givord, unpublished data) produces local lesions in C. amaranticolor, N. tabacum cv. Samsun and N. glutinosa, systemically infects C. sativus cv. Vert Long Maraîcher and P. vulgaris cv. The Prince, and is not transmissible to V. rosea. A yellow mosaic is produced in okra.
Transmission by Vectors
Not soil-borne or transmitted by Aphis gossypii or Bemisia tabaci (Givord et al., 1972); beetles have not yet been tested as vectors.
Transmission through Seed
Not seed-borne in okra.
Transmission by Dodder
No transmission by Cuscuta subinclusa (Givord & Hirth, 1973).
Serology
Strongly immunogenic in rabbits. Antisera with precipitin titres of 1/1000 are readily obtained. In gel double diffusion tests the virus gives one line of precipitate.
Relationships
The virus belongs to the tymovirus group and has serological relationships with Andean potato latent, belladonna mottle, cacao yellow mosaic, desmodium yellow mottle, dulcamara mottle, eggplant mosaic, ononis yellow mosaic, scrophularia mottle, turnip yellow mosaic and wild cucumber mosaic viruses. Most of these relationships are rather distant, but that to desmodium yellow mottle virus is closer (Koenig & Givord, 1974). The virus has many properties in common with viruses in the tymovirus group, e.g. sedimentation as two components (one consisting of empty protein shells), high concentration in host plants, high thermal inactivation point, high cytidilic acid content of the RNA.
Stability in Sap
In okra sap, the thermal inactivation point (10 min) is about 80°C, dilution end-point about 10-6, and infectivity is retained at 24°C for 7-9 days (Givord et al., 1972).
Purification
The virus is readily purified by one of the following methods:
1. The bentonite/n-butanol method of Givord & Hirth (1973). Mince 100 g infected okra leaves with 150 ml 0.01 M sodium phosphate buffer, pH 7, containing 0.5% sodium bisulphite and 1% bentonite. Centrifuge expressed sap at 12,000 g and emulsify supernatant fluid with 0.5 volume of n-butanol. Keep mixture for 1 h at 4°C and centrifuge at low speed. Bubble nitrogen through aqueous phase and separate the virus by three cycles of differential centrifugation, re-suspending pellets in 0.1 M sodium phosphate (pH 7.3) containing 0.01 M EDTA.
2. The magnesium bentonite method of Dunn & Hitchborn (1965), as recommended for turnip yellow mosaic virus. Mince 100 g infected cucumber leaves with 5 ml 0.1 M magnesium sulphate and 50 ml of a magnesium bentonite suspension (50 mg magnesium bentonite per 1 ml 0.001 M phosphate buffer, pH 7.4, containing 0.001 M magnesium sulphate; for details of preparation see Dunn & Hitchborn, 1965). Centrifuge the expressed sap at 15,000 g and separate the virus from the supernatant fluid by high speed centrifugation. Resuspend pellets in 10 ml 0.01 M phosphate buffer, pH 7.4, containing 0.005 M magnesium sulphate and 1 ml of the magnesium bentonite suspension. Repeat differential centrifugation and resuspend the virus in dilute buffer.
Properties of Particles
The particles are all the same size, but sediment as two components (Fig. 7), protein shells without nucleic acid (T), and infective nucleoprotein (B).
Sedimentation coefficient (s20): 42 S (T) and 106 S (B) in phosphate buffer (0.01 M, pH 7.0) at 22.7°C. Calculation was not made for infinite dilution.
Electrophoretic behaviour: in immunoelectrophoresis in 1% agarose containing 0.025 M Tris-HCl buffer, pH 8, the virus migrates towards the cathode (R. Koenig, unpublished).
Absorbance at 260 nm (determined by differential refractometry in the analytical ultracentrifuge): about 9 for the normal mixture of the two components at 1 mg/ml with a light path of 1 cm (J. P. Bouley and B. Dorne, unpublished data).
A260/A280: 1.61 (B).
Particle Structure
Particles are isometric c. 28 nm in diameter (Fig. 8) with icosahedral symmetry and 32 morphological subunits. T particles are penetrated by 1% uranyl acetate (Givord & Hirth, 1973). In structure the virus apparently resembles other tymoviruses.
Particle Composition
Nucleic acid: RNA, single-stranded, 32% of the particle weight (calculated according to Reichmann, 1965; J. P. Bouley & B. Dorne, unpublished data). Molar percentage of nucleotides: G17.2; A17.5; C39.8; U25.5 (Givord & Hirth, 1973).
Protein: M. Wt 20,000 by SDS-polyacrylamide electrophoresis, using 10% acrylamide (Givord & Hirth, 1973).
Relations with Cells and Tissues
No information.
Notes
Okra yellow vein virus may cause rather similar symptoms in okra but is transmissible by white flies (Aleyrodidae) and not mechanically. Some other diseases are found in okra and Hibiscus rosa-sinensis but they have very different symptoms and their causal agents are not readily transmitted by inoculation of sap: Hibiscus esculentus vein-thickening, H. esculentus leaf curl, H. rosa-sinensis line pattern, H. rosa-sinensis leaf curl. For details of viruses and virus-like agents that can infect H. esculentus or other malvaceous plants, see Givord & Hirth (1973).
Figures

Systemically infected okra leaf (Hibiscus esculentus cv. Clemson spineless). (Photo: G. Petiot.)

Systemically infected okra leaf (Hibiscus esculentus cv. Clemson spineless). (Photo: G. Petiot.)

Systemically infected okra leaf (Hibiscus esculentus cv. Clemson spineless). (Photo: G. Petiot.)

Systemically infected okra leaf (Hibiscus esculentus cv. Clemson spineless). (Photo: G. Petiot.)

Systemically infected cowpea leaf. (Photo: L. Givord.)

Systemically infected groundnut leaf. (Photo: L. Givord.)

Schlieren pattern obtained by analytical ultracentrifugation of a purified virus preparation showing top and bottom components. Sedimentation is from left to right.
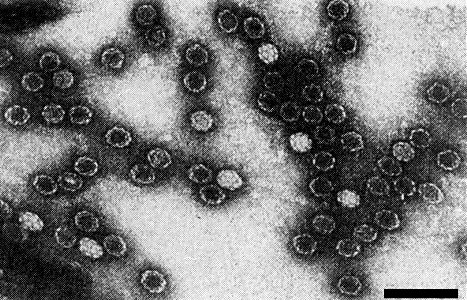
Virus particles over a hole in the carbon film. Uranyl acetate staining. Bar represents 100 nm. (Photo: P. Pfeiffer.)
References list for DPV: Okra mosaic virus (128)
- Dunn & Hitchborn, Virology 25: 171, 1965.
- Givord & Hirth, Ann. appl. Biol. 74: 359, 1973.
- Givord, Pfeiffer & Hirth, C.r. hebd. Séanc. Acad. Sci., Paris 275: 1563, 1972.
- Koenig & Givord, Virology 58: 119, 1974.
- Reichmann, Virology 25: 166, 1965.