Details of DPV and References
DPV NO: 131 July 1974
Family: Potyviridae
Genus: Potyvirus
Species: Clover yellow vein virus | Acronym: ClYVV
Clover yellow vein virus
M. Hollings Glasshouse Crops Research Institute, Littlehampton, Sussex, England
Olwen M. Stone Glasshouse Crops Research Institute, Littlehampton, Sussex, England
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
- Described by Hollings & Nariani (1965).
- An RNA-containing virus with filamentous particles c. 760 nm long, which infects several species in the Leguminosae (especially Trifolium spp.) and in five other families. It is readily transmitted by inoculation of sap, and by aphids in the non-persistent manner.
Main Diseases
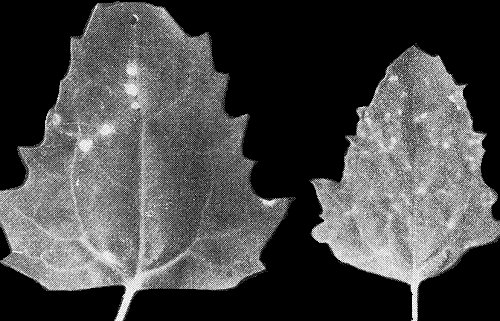
Causes mild veinal yellowing and mottle in Trifolium spp. (Fig. 5), especially in white clover (T. repens); induces vein-clearing and mosaic in coriander (Coriandrum sativum).
Geographical Distribution
Recorded in Britain (Hollings & Nariani, 1965 Gibbs,Varma & Woods, 1966), and in Canada (Pratt, 1968; Singh & Lopez-Abella, 1971); it may also occur in the USA (Pratt, 1969).
Host Range and Symptomatology
Known hosts comprise about 25 species in 6 plant families.
- Diagnostic species
- No single test plant gives diagnostic reactions.
- Chenopodium amaranticolor. Necrotic local lesions 1-2 mm diameter after c. 7 days, enlarging to form necrotic areas, and followed by abscission of the inoculated leaves. No systemic infection.
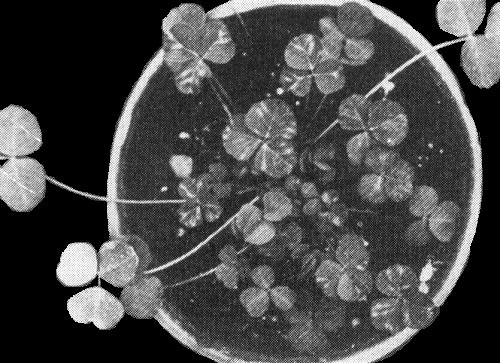
- C. quinoa. Fawn, necrotic local lesions 1-2 mm diameter after 6-9 days; systemic chlorosis, flecks and necrotic spots develop in 2 weeks (Fig. 1).
- Nicotiana clevelandii. Occasionally, local chlorotic lesions after 6 days; systemic chlorotic speckles and mottle after 1-3 weeks, intensifying with time. Plants are dwarfed (Fig. 4).
- N. tabacum (tobacco), White Burley cv. Judy’s Pride. Chlorotic local lesions after about 6 days; no systemic infection.
- Phaseolus vulgaris (French bean) cv. The Prince. Few chlorotic local lesions in about 7 days. Systemic chlorotic flecks or vein-clearing in the younger leaves 10-14 days after infection; later a granular yellow mosaic, sometimes with larger irregular chlorotic or pale necrotic areas in the older leaves (Fig. 3).
- Tetragonia expansa. Fawn, semi-necrotic local lesions c. 3 mm diameter after 7-12 days: lesions later develop necrotic margins and often form broken rings (Fig. 2).
- The virus does not infect Vigna sinensis; some isolates infect certain cultivars of Pisum sativum and Vicia faba.
- Chenopodium amaranticolor. Necrotic local lesions 1-2 mm diameter after c. 7 days, enlarging to form necrotic areas, and followed by abscission of the inoculated leaves. No systemic infection.
- Propagation species
- Nicotiana clevelandii is a good source of virus for purification, and for
maintaining virus cultures.
- Assay species
- Chenopodium quinoa
gives satisfactory local lesions and is more sensitive than C. amaranticolor.
Strains
The type strain was obtained from Trifolium repens in Britain (Hollings & Nariani, 1965); other isolates from Trifolium have shown minor differences in host range. An isolate infecting coriander (Singh & Lopez-Abella, 1971) was less stable in vitro than the type strain, and induced systemic vein-clearing, mosaic and eventual wilt in pea (Pisum sativum), whereas the type strain did not infect pea. The type strain readily infected coriander, and antiserum to type strain reacted to titre with the coriander isolate.
Transmission by Vectors
Transmitted in the non-persistent manner by the aphids Myzus persicae, Acyrthosiphon pisum, Aulacorthum solani and Macrosiphum euphorbiae, but not by Aphis fabae (Hollings & Nariani, 1965; Singh & Lopez-Abella, 1971).
Transmission through Seed
No transmission through seed of Phaseolus vulgaris cv. The Prince (Hollings & Nariani, 1965).
Transmission by Dodder
No information.
Serology
The virus is a good immunogen, although it is difficult to obtain virus preparations completely free from contaminating plant proteins. Rabbits immunized with one intravenous plus two intramuscular injections (with Freund’s complete adjuvant) gave antisera with specific titres in precipitin tubes of 1/16,384 but titres of c. 1/2000 to healthy plant antigens. However, antisera obtained by immunizing rabbits with virus preparations clarified with n-butanol (which fragments the particles) reacted well against intact virus and contained very few antibodies to plant material (Hollings & Nariani, 1965). Precipitates are flocculent (flagellar) in tube and in microprecipitin tests; ring tests are also satisfactory. Gel-diffusion tests are only possible with fragmented virus; best results were obtained with 0.8% Ionagar containing 0.2 M (NH4)2HPO4 +1% ethanolamine + 0.2% Tween 20.
Relationships
Distantly related serologically to bean common mosaic, bean yellow mosaic, soybean mosaic, sugarcane mosaic and turnip mosaic viruses (Gibbs et al., 1966), and to carnation vein mottle, pepper veinal mottle, and un-named viruses from freesia, and from sweet potato in New Zealand (Hollings & Stone, 1973).
Stability in Sap
In Nicotiana clevelandii sap, the thermal inactivation point (10 min) is 55-60°C (coriander isolate 50-55°C), the dilution end-point is 10-4 to 10-5, and infectivity is retained for up to 8 days at c. 18°C and for at least 11 weeks at 0°C. Lyophilized sap (with 7%, peptone + 7% dextrose) retains infectivity in vacuo at laboratory temperature for at least 9 years.
Purification
The virus withstands clarification of sap with chloroform, diethyl ether or carbon tetrachloride, but the particles are fragmented by n-butanol. Yields of 10-20 mg virus can be obtained from 1 kg leaf tissue of Nicotiana clevelandii, using either of the following methods, but a density-gradient centrifugation stage is necessary to obtain pure preparations.
1. (Hollings & Nariani, 1965). Harvest Nicotiana clevelandii plants c. 2 weeks after infection and chill the leaves, then homogenize each 100 g at 0°C with 200 ml of 0.1 M phosphate-ascorbic acid buffer (pH 7.0) containing 0.1% thioglycollic acid + 25 ml diethylether and 25 ml carbon tetrachloride. Shake the chilled mixture for 15 min, express the juice through cheesecloth and separate the emulsion by low-speed centrifugation. Sediment the virus by one cycle of differential centrifugation. In further purification by sucrose density-gradient centrifugation, dilute the extracted virus-containing band with 0.5 M phosphate buffer and re-sediment the virus by centrifugation.
2. (Modified from Brunt & Kenten, 1971). Harvest the N. clevelandii plants as above, homogenize at room temperature with 0.5 M borate buffer (pH 7.8) containing 0.01 M MgCl2, and 0.2% 2-mercaptoethanol, using 2 ml buffer/g leaf. Add one-half volume of CHCl3, again homogenize, and allow to stand for 30 min. Clarify by low-speed centrifugation and filter the aqueous phase (Whatman No. 4 paper). Add polyethylene glycol (PEG 6000 M. Wt, 5 g/100 ml of fluid) and keep for 90 min at 2°C. Sediment the precipitate (10 min, 10,000 g). Re-suspend the precipitate in about one-tenth original volume of extracting buffer and allow to stand overnight at c. 2°C. Add extracting buffer up to the original volume and concentrate the virus by one cycle of differential centrifugation (30 min at 10,000 g; 90 min at 78,000 g). Further purify by sucrose density-gradient centrifugation.
Properties of Particles
Sedimentation coefficient (s°20, w): 159 S. No accessory particles are found by analytical ultracentrifugation.
A260/A280: 1.29: Amax(260)/Amin(246): 1.18 (values corrected for light-scattering).
Particle Structure
Particles are filamentous and flexuous with a modal length of 767 nm (Gibbs et al., 1966) x 12 nm in width; many particles appear shorter than this (590-700 nm) (Hollings & Nariani, 1965; Singh & Lopez-Abella, 1971), probably because they fragment easily (Fig. 6). When mounted in uranyl formate, the surface of the particles appears verrucose, with a mean pitch of the basic helix of 35 Å (Varma et al., 1968).
Particle length is affected by the presence of magnesium ions. In 0.05 M MgCl2, the particles are almost straight and c. 900 nm long. In 0.02 M di-sodium ethylenediamine-tetraacetate (EDTA) at pH 7.2 they are very slightly flexuous and c. 830 nm long; in 0.03 M EDTA (pH 7.4) they are c. 800 nm long; and in 0.1 M EDTA (pH 6.4) they are markedly more flexuous and c. 760 nm long (M. Hollings & O. M. Stone, unpublished results).
Particle Composition
Nucleic acid: Preliminary results suggest the virus contains single-stranded RNA (M. Hollings & O. M. Stone, unpublished results).
Protein: Protein from purified virus preparations usually produces three peaks in polyacrylamide gel electrophoresis, corresponding to M. Wt 29,900, 31,600 and 34,600, but preparations obtained from density-gradients containing 0.1% diethyl pyrocarbonate gave only one protein, of M. Wt 34,400 (R. J. Barton, M. Hollings & O. M. Stone, unpublished results).
Relations with Cells and Tissues
Amorphous, granular intracellular inclusion bodies are present in epidermal cells of infected Nicotiana clevelandii (Hollings & Nariani, 1965) and of Vicia faba and Pisum sativum (Bos, 1969). In ultrathin sections, pinwheel and bundle inclusions are found, similar to those induced by other members of the potyvirus group (Pratt, 1969). The virus also causes irregular enlargement of the nucleoli (Bos, 1969).
Notes
Clover yellow vein virus is a typical member of the potyvirus group. It is most reliably distinguished from other members of the group found in Leguminosae (bean mosaic, bean yellow mosaic, pea mosaic, lupin mottle and soybean mosaic viruses) by serological tests, because there are no consistent differences in host ranges and symptoms among the many described strains of these viruses. Red clover vein mosaic and pea streak viruses (carlavirus group) induce local lesions in Gomphrena globosa, whereas clover yellow vein virus does not. The virus is most easily differentiated from the many isometric viruses of Leguminosae by electron microscopy. Clover yellow vein virus was not eliminated from Nicotiana clevelandii plants grown for 4 weeks at 38°C.
Figures
Local lesions (left) and systemic chlorotic flecks and mottle (right) in Chenopodium quinoa.

Local lesions in Tetragonia expansa 18 days after inoculation.

Systemic mottle and slight distortion in Phaseolus vulgaris cv. The Prince 3 weeks after inoculation.

Plant of Nicotiana clevelandii 6 weeks after infection, showing systemic chlorotic mottle, some leaf distortion, and stunting.
Systemic vein yellowing and mottle in Trifolium incarnatum 18 days after infection.

Virus particles in partially purified preparation mounted in potassium phosphotungstate. Bar represents 500 nm.
References list for DPV: Clover yellow vein virus (131)
- Bos, Neth. J. Pl. Path. 75:137, 1969.
- Brunt & Kenten, Ann. appl. Biol. 69: 235, 1971.
- Gibbs, Varma & Woods, Ann. appl. Biol. 58: 231, 1966.
- Hollings & Nariani, Ann. appl. Biol. 56: 99, 1965.
- Hollings & Stone, Rep. Glasshouse Crops Res. Inst. 1972: 107, 1973.
- Pratt, Can. Pl. Dis. Surv. 48: 87, 1968.
- Pratt, Pl. Dis. Reptr 53: 210, 1969.
- Singh & Lopez-Abella, Phytopathology 61: 333, 1971.
- Varma, Gibbs, Woods & Finch, J. gen. Virol. 2: 107, 1968.