Details of DPV and References
DPV NO: 141 October 1975
Family: Potyviridae
Genus: Potyvirus
Species: Peanut mottle virus | Acronym: PeMoV
Peanut mottle virus
K. R. Bock Overseas Development Administration, Crop Virology Project, EAAFRO P.O. Box 30148, Nairobi, Kenya
C. W. Kuhn Department of Plant Pathology and Plant Genetics, University of Georgia, Athens, Georgia 30602, USA
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
- Described by Kuhn (1965) and Bock (1973).
- Synonyms
- Peanut mild mosaic virus (Rev. appl. Mycol. 29: 397)
- Peanut severe mosaic virus (Rev. appl. Mycol. 29: 397)
- Groundnut mottle virus (Rev. appl. Mycol. 47: 2420)
- Peanut severe mosaic virus (Rev. appl. Mycol. 29: 397)
- A serologically distinct RNA-containing virus of the potyvirus group with flexuous filamentous particles c. 740-750 nm long. It has a restricted experimental host range, is transmitted by inoculation of sap and by several common species of aphid, is seed-borne in groundnut, and probably occurs wherever groundnuts are grown.
Main Diseases
Causes systemic mottle and necrosis in groundnut (Arachis hypogaea), bean (Phaseolus vulgaris), soybean (Glycine max) and pea (Pisum sativum), the severity of disease being dependent on host cultivar and virus strain. Economically significant in groundnut in USA (where Kuhn (1965) reported a yield loss of 25% in greenhouse tests), and in bean in Australia (Behncken & McCarthy, 1973).
Geographical Distribution
Occurs in south-east USA, East Africa and north-east Australia, and probably in Japan, West Malaysia, South America (Venezuela) and Europe (Bulgaria).
Host Range and Symptomatology
Although the virus infects several species within the Leguminosae, its host range outside this family is extremely limited. Readily transmissible by inoculation of sap.
- Diagnostic species
- Phaseolus vulgaris
(French bean). Local necrotic lesions which spread dendritically along veins (Fig. 1). No systemic infection in many cvs. (e.g. The Prince, Topcrop, Tendergreen). In others (e.g. California Small White, Gallaroy, Pinto), systemic mottle with limited or severe necrosis (Behncken, 1970; Behncken & McCarthy, 1973). - Arachis hypogaea (groundnut). The most commonly occurring and widespread
(mild) strains induce a mild dark-green mottle (Fig. 2); leaflet margins are
crinkled with depression of interveinal tissue. For symptoms induced by other
isolates, see ‘Strains’.
- Glycine max (soybean). Chlorotic or necrotic local lesions with some cvs; systemic chlorotic mottle, mild mosaic or mild to severe leaf-curling and distortion with others (Fig. 5).
- Cassia occidentalis. Systemic yellow mottle induced by most strains (Fig. 6), but the severe strain (S) does not infect.
- Nicotiana clevelandii. Chlorotic or necrotic local lesions; systemic mottle.
- Chenopodium amaranticolor. Apparently immune (Kuhn, 1965; Behncken, 1970; Sun & Hebert, 1972; Bock, 1973).
- Glycine max (soybean). Chlorotic or necrotic local lesions with some cvs; systemic chlorotic mottle, mild mosaic or mild to severe leaf-curling and distortion with others (Fig. 5).
- Propagation species
- Cultures are best maintained in Cassia occidentalis or groundnut
(for severe strain). For purification the virus should be propagated in suitable
cvs. of pea, cowpea or bean.
- Assay species
- Phaseolus vulgaris
(cv. The Prince, Tendergreen or Topcrop) is a good local lesion host.
Strains
The American mild (type) strain (M-2) (Paguio & Kuhn, 1973a), the East African groundnut and soybean isolates, and an Australian isolate from bean are serologically indistinguishable from each other (K. R. Bock & G. W. Behncken, unpublished data) and closely resemble each other in the symptoms they induce in groundnut.
At present there is no evidence for different strains in Africa and Australia. In USA, however, Paguio & Kuhn (1973a) have differentiated five serologically indistinguishable variants, chiefly on symptom expression in groundnut: M-1 and M-2 (mild mottle); N (necrosis, Fig. 3); S (severe mosaic = severe strain of Sun & Hebert, 1972; Fig. 4); and CLP (chlorotic line pattern, Fig. 7). The M-2 strain cross-protects against N, S, and CLP in groundnut.
Peanut mild mosaic and peanut severe mosaic viruses (Cooper, 1950) are synonymous with peanut mottle virus M and S strains, respectively (Sun & Hebert, 1972).
Transmission by Vectors
The virus is transmitted in the non-persistent manner by the aphids Aphis craccivora, A. gossypii, Hyperomyzus lactucae, Myzus persicae and Rhopalosiphum padi (Behncken, 1970). A. craccivora and M. persicae can transmit strains M1, M2, S, and CLP but not strain N. However, both aphids can transmit strain N if it is in a plant simultaneously inoculated with strain M2 (O. R. Paguio & C. W. Kuhn, unpublished data).
Transmission through Seed
Seed-transmission occurs in groundnut at low rates (0.02-2.0%) but apparently not in cowpea, soybean, Pisum sativum, or Cassia obtusifolia (Kuhn, 1965; Paguio & Kuhn, 1973a; C. W. Kuhn, unpublished data). Seed-transmission (<1%) has been reported also in bean (Behncken & McCarthy, 1973). Twenty percent of the groundnut plants developing from pods left in situ in pots were infected with virus (Bock, 1973).
Transmission by Dodder
Not reported.
Serology
The virus is moderately to strongly immunogenic; an antiserum with a tube precipitin titre of 1/8192 has been obtained (Bock, 1973).
Relationships
Peanut mottle virus does not react with antisera to the following common potyviruses: bean common mosaic, bean yellow mosaic, cowpea aphid-borne mosaic, soybean mosaic, clover yellow vein, potato Y, tobacco severe etch, celery mosaic, iris mosaic and sugarcane mosaic (Bock, 1973; Sun & Hebert, 1972).
In East Africa, viruses serologically related to peanut mottle have been obtained from Cassia (K. R. Bock, unpublished data) and Voandzeia (Bock, 1973).
The Venezuelan (Herold & Munz, 1969), Japanese (Inouye, 1969) and European (Schmidt & Schmelzer, 1966) viruses have not been tested serologically against antisera to American, East African or Australian isolates, but they closely resemble peanut mottle virus in host range and symptomatology, properties in vitro, transmission by aphids and particle morphology.
Groundnut mosaic virus (Ting, Geh & Chuan, 1972), found in West Malaysia, is probably the same as or closely related to peanut mottle virus. The two viruses share many properties but particle characteristics and serological reactions have not been determined for the Malaysian virus.
Stability in Sap
The thermal inactivation point of the East African and Australian isolates is between 55° and 59°C; that of the American isolates is 60 to 64°C. The dilution end-point of all isolates is between 10-3 and 10-4, and infectivity is retained at 20°C for 1-2 days (but less than 1 day with the S strain). Frozen infected leaves retain infectivity for at least 12 wk at -12°C (Bock, 1973).
Purification
Two methods appear to be satisfactory.
1. Bock (1973). Homogenize systemically infected cowpea leaves at pH 8.1 in 0.5 M sodium citrate buffer containing 1%, mercaptoethanol, (1 ml/g tissue), clarify by treatment with equal volume of chloroform; centrifuge differentially; resuspend in 0.01 M phosphate buffer, pH 7.7.
2. Paguio & Kuhn (1973b). Homogenize systemically infected pea leaves at pH 8 in 0.01 M potassium phosphate buffer, containing 0.01 M DIECA, 0.01 M sodium bisulphite (2 ml/g tissue), clarify by adding 0.1 vol chloroform and centrifuging at 10,000 g for 10 min. Precipitate virus by adding polyethylene glycol (PEG) to 4% and KCl to 0.2 M; centrifuge at low speed and resuspend pellets in 0.05 M phosphate buffer containing 0.001 M dithiothreitol. Repeat the PEG-KCl precipitation and follow with rate zonal density-gradient centrifugation.
Properties of Particles
A260/A280: 1.24 (Paguio & Kuhn, 1973b).
A260(max)/A246 (min): 1.3 (Paguio & Kuhn, 1973b).
Absorbance at 260 nm (1 mg/ml, 1 cm light path): 2.6 (C. W. Kuhn, unpublished data).
Particle Structure
Particles are flexuous filaments (Fig. 8), probably c. 740-750 nm long (Sun & Hebert, 1972; Bock, 1973; Paguio & Kuhn, 1973b), but estimates of length range between 704 nm (Behncken, 1970) and 812 nm (Herold & Munz, 1969).
Particle Composition
Nucleic acid: RNA, probably single-stranded, about 6% of the particle weight (C. W. Kuhn, unpublished data).
Protein: No information.
Relations with Cells and Tissues
Virus particles occur in the cytoplasm singly, or in loosely arranged bundles. Pinwheel inclusions and plates typical of the potyvirus group are seen in tissue sections (Herold & Munz, 1969; Sun & Hebert, 1972).
Notes
The following viruses important in field-grown groundnut have been described sufficiently well to permit comparison with peanut mottle virus. Tomato spotted wilt virus (Ie, 1970) is easily differentiated from peanut mottle virus by its low thermal inactivation point (40-46°C), its instability in sap and its very wide host range. The two component viruses of groundnut rosette disease are persistent in Aphis craccivora (Storey & Ryland, 1955); peanut stunt virus (Mink, 1972) has isometric particles and a wide host range; cowpea mild mottle virus (Brunt, 1974) has particles 650 nm long and is not transmitted by aphids.
Figures
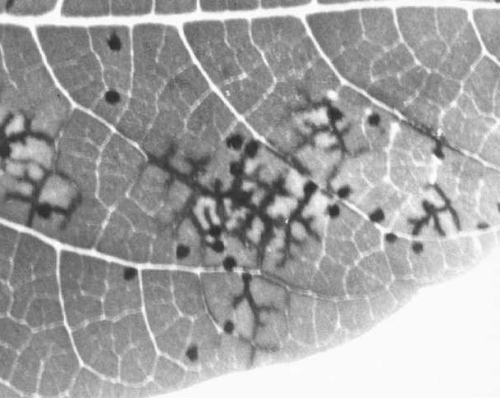
Local lesions induced in Phaseolus vulgaris (cv. Topcrop ) by type (M-2) strains.

Systemic symptoms induced in groundnut (cv. Argentine) by mild (M-2) strain.

Systemic symptoms induced in groundnut (cv. Argentine) by necrosis (N) strain.

Systemic symptoms induced in groundnut (cv. Argentine) by severe (S) strain.

Systemic symptoms induced in soybean (cv. HLS 541) by East African isolate.

Systemic symptoms induced in Cassia occidentalis by East African isolate.

Systemic symptoms induced in groundnut (cv. Argentine) by chlorotic line pattern (CLP) strain.

Virus particles from a purified preparation of an African isolate in phosphotungstate. Bar represents 200 nm.
References list for DPV: Peanut mottle virus (141)
- Behncken, Aust. J. agric. Res. 21: 465, 1970.
- Behncken & McCarthy, Qd. agric. J. 99: 635, 1973.
- Bock, Ann. appl. Biol. 74: 171, 1973.
- Brunt, CMI/AAB Descriptions of Plant Viruses 140, 4 pp., 1974.
- Cooper, Phytopathology 40: 6, 1950.
- Herold & Munz, Phytopathology 59: 663, 1969.
- Ie, CMI/AAB Descriptions of Plant Viruses 39, 4pp., 1970.
- Inouye, Nogaku Kenkyu 52: 159, 1969.
- Kuhn, Phytopathology 55: 880, 1965.
- Mink, CMI/AAB Descriptions of Plant Viruses 92, 4 pp., 1972.
- Paguio & Kuhn, Phytopathology 63: 976, 1973a.
- Paguio & Kuhn, Phytopathology 63: 720, 1973b.
- Schmidt & Schmelzer, Phytopath. Z. 55: 92, 1966.
- Storey & Ryland, Ann. appl. Biol. 43: 423, 1955.
- Sun & Hebert, Phytopathology 62: 832, 1972.
- Ting, Geh & Chuan, Expl Agric. 8: 355, 1972.