Details of DPV and References
DPV NO: 152 October 1975
Family: Virgaviridae
Genus: Tobamovirus
Species: Ribgrass mosaic virus | Acronym: RMV
Ribgrass mosaic virus
N. Oshima Institute for Plant Virus Research, Chiba, Chiba-Ken, Japan
B. D. Harrison Scottish Horticultural Research Institute, Invergowrie, Dundee, Scotland
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
Described by
Holmes (1941).
Selected synonyms
- Holmes’ ribgrass virus
- Marmor tabaci var. plantaginis (Rev. appl. Mycol. 21: 227)
- Plantago mosaic virus (Rev. appl. Mycol. 21: 227)
- Plantago virus (Rev. appl. Mycol. 38: 338)
- Ribgrass strain of tobacco mosaic virus (Rev. appl. Mycol. 21: 227)
- Marmor tabaci var. plantaginis (Rev. appl. Mycol. 21: 227)
-
An RNA-containing virus with rod-shaped particles, 18 nm wide and 300 nm long. It is found in several wild plant species, occurs in high concentration in sap, is very stable and is readily transmitted by inoculation of sap. No vector known.
Main Diseases
Causes systemic chlorotic mottling and ring-like markings, chlorotic streaks along the veins and slight twisting of the petioles in ribgrass (Plantago lanceolata) (Fig. 1) and broad-leaved plantain (P. major), mosaic in Digitalis lanata, mosaic/mottle in Melandrium album (Kovachevsky, 1963), mottle in various cruciferous species, necrotic mosaic in tobacco (Valleau & Johnson, 1943), and internal browning of the fruits of tomato (Holmes, 1950).
Geographical Distribution
Widely distributed. Reported in Asia, Europe, N. America and USSR.
Host Range and Symptomatology
Hosts infected by inoculation of sap include about 50 species in about 10 plant families, especially in the Solanaceae, Plantaginaceae, Cruciferae and Scrophulariaceae (Kovachevsky, 1963). Symptoms are mostly mosaics and mottles, but several cruciferous species may be infected symptomlessly (Oshima et al., 1974).
-
Diagnostic species
- Chenopodium amaranticolor.
Small necrotic local lesions. Uninoculated tip leaves may develop distortion with necrotic spots. - Cyphomandra betacea. Chlorotic local lesions. Systemic distorting mosaic.
- Nicotiana glutinosa, N. sylvestris and N. tabacum cv. Xanthi-nc. Small brown necrotic local lesions. No systemic infection.
- Nicotiana tabacum (tobacco). Turkish tobacco develops local lesions as thin, white rings of necrotic tissue in inoculated leaves. Systemic symptoms consist of rings and lines of necrotic tissue along the veins. Both local and systemic symptoms are sometimes zonate (Fig. 3). White Burley tobacco develops chlorotic local lesions but is seldom infected systemically.
- Phaseolus vulgaris (bean). Not infected.
- Nicotiana glutinosa, N. sylvestris and N. tabacum cv. Xanthi-nc. Small brown necrotic local lesions. No systemic infection.
-
Propagation species
- N. tabacum
cv. Samsun.Assay species
- N. tabacum
cv. Xanthi-nc and N. glutinosa.
Strains
Many minor variants have been isolated from various species:
Type Strain (Holmes, 1941). Obtained from Plantago lanceolata in New Jersey, USA.
Digitalis isolate (Schumann, 1963). Obtained from Digitalis lanata in Germany.
Lychnis isolate (Chessin et al., 1967). Obtained from Melandrium album (Lychnis alba) in Montana, USA.
Necrotic isolate (Polák, 1962). Obtained from Cordaria draba, Sisymbrium loeselii and Anchusa officinalis in Czechoslovakia.
Primula isolate (Kovachevsky, 1969). Obtained from Primula obconica in Bulgaria.
RMV-K (Juretic, Milicic & Mamula, 1973). Obtained from Rorippa amphibia in Yugoslavia.
TMV-C (Goto & Oshima, 1962). Obtained from Radicula sylvestris in northern Japan (Fig. 2).
Wasabi isolate (Tochihara et al., 1964). Obtained from Eutrena wasabi in central Japan. In polyacrylamide gels containing urea, the protein of this strain migrates faster during electrophoresis than do those of TMV-C, ribgrass mosaic type strain, and lychnis isolates (Fig. 7) (Oshima et al., 1971).
Youcai mosaic isolate (YMV15) (Pei, 1962). Apparently similar to ribgrass mosaic type strain. Obtained from Brassica campestris in China.
Transmission by Vectors
None found.
Transmission through Seed
None found.
Transmission by Dodder
None found.
Serology
Antisera with titres in tube precipitin tests of 1/1000 are readily obtained. Double diffusion tests can be made in agar gel (e.g. Juretic et al., 1969).
Relationships
Ribgrass mosaic virus strains are more closely related serologically to tobacco mosaic (TMV) and tomato mosaic viruses than to other tobamoviruses (van Regenmortel, 1967; Hollings, 1974). Tobacco plants infected with the type strain of TMV do not develop additional symptoms after inoculation with the ribgrass strain (Holmes, 1941).
Among the minor variants listed under ‘Strains’, the necrotic and Primula isolates are serologically similar, but distinguishable from the Digitalis and RMV-K isolates, which themselves are probably serologically similar; all differ slightly from the type isolate of the ribgrass strain (Juretic & Wetter, 1973; Juretic, 1974).
Stability in Sap
In infected Turkish tobacco sap, the thermal inactivation point (10 min) is 93°C, the dilution end-point 10-6-10-7. The virus is infective after more than 5 months in sap, and more than 122 days in dried tobacco leaves (Holmes, 1941).
Purification
The virus is purified from extracts of systemically infected leaves of Samsun tobacco by freezing, followed by thawing and differential centrifugation. Several methods used for the type strain of TMV can also be used. Purified virus lacks the brown colour typical of type strain TMV, which is caused by plant material binding to the particles (Siegel & Wildman, 1954).
Properties of Particles
Molecular weight: About 40 x 106.
Isoelectric point: pH 4.49 (Oster, 1951).
Electrophoretic mobility: -7.7 x 10-5 cm2/sec/volt at pH 7.1 in 0.05 M phosphate buffer (Knight & Lauffer, 1942).
Absorbance at 260 nm (1 mg/ml, 1 cm light path): Probably the same as type strain TMV (3.2).
A260/A280: 1.2.
Inactivation by ultraviolet irradiation: More sensitive than type strain TMV, slightly less sensitive than strain U2 (Siegel & Wildman, 1954).
Particle Structure
The particles are straight rods, 18 nm wide and 300 nm long with ends not rounded (Fig. 4). The rods are helically constructed with a pitch of about 2.3 nm (Fig. 5); the RNA is embedded in the helical array of identical coat protein subunits. Particles have a central channel.
Particle Composition
Nucleic acid: Single-stranded RNA, 5% of particle weight, M. Wt about 2 x 106. Molar percentage of nucleotides: G25.8; A29.3; C18.0; U27.0 (Knight, 1952).
Protein: 95% of particle weight, composed of about 2000 chemical subunits per particle, each of M. Wt about 17,500.
The coat protein consists of 156 amino acids whose sequence is as follows (Wittmann et al., 1969):
10
AcSer -Tyr-Asn-Ile-Thr-Asn-Ser-Asn-Gln-
Tyr-Gln-Tyr-Phe-Ala-Ala-Val-Trp-Ala-Glu-
20 30
Pro-Thr-Pro-Met-Leu-Asn-Gln-Cys-Val-Ser-
Ala-Leu-Ser-Gln-Ser-Tyr-Gln-Thr-Gln-Ala-
40 50
Gly-Arg-Asp-Thr-Val-Arg-Gln-Gln-Phe-Ala-
Asn-Leu-Leu-Ser-Thr-Ile-Val-Ala-Pro-Asn-
60 70
Gln-Arg-Phe-Pro-Asp-Thr-Gly-Phe-Arg-Val-
Tyr-Val-Asn-Ser-Ala-Val-Ile-Lys-Pro-Leu-
80 90
Tyr-Glu-Ala-Leu-Met-Lys-Ser-Phe-Asp-Thr-
Arg-Asn-Arg-Ile-Ile-Gln-Thr-Glu-Glu-Gln-
100 110
Ser-Arg-Pro-Ser-Ala-Ser-Gln-Val-Ala-Asn-
Ala-Thr-Gln-Arg-Val-Asp-Asp-Ala-Thr-Val-
120 130
Ala-Ile-Arg-Ser-Gln-Ile-Gln-Leu-Leu-Leu-
Asn-Glu-Leu-Ser-Asn-His-Gly-Gly-Tyr-Met-
140 150
Asn-Arg-Ala-Glu-Phe-Glu-Ala-Ile-Leu-Pro-
Trp-Thr-Thr-Ala-Pro-Ala-Thr.
The protein is noteworthy for containing histidine and methionine.
Relations with Cells and Tissues
Several isolates induce characteristic crystalline inclusions in infected cells, the so-called ‘rounded plates’. These are discs of various sizes, each consisting of one or more layers containing virus particles orientated perpendicularly to the surface of the plates (Goldin, 1953; Milicic et al., 1968; Fig. 6). The RMV-K isolate induces X-bodies, but not rounded plates (Juretic, 1974).
Notes
Ribgrass mosaic virus can be distinguished from other viruses infecting Plantago by its particle shape and high thermal inactivation point. It is distinguished from other tobamoviruses serologically, by the symptoms produced in diagnostic hosts, and by its consistent ability to infect systemically both Plantago lanceolata and P. major.
Figures

Leaves of Plantago lanceolata systemically infected with ribgrass mosaic type strain.

Tobacco plant cv. Samsun systemically infected with the TMV-C isolate.

Leaf of Nicotiana tabacum cv. Samsun systemically infected with ribgrass mosaic type strain, showing fine necrotic line patterns.

Virus particles of ribgrass mosaic type strain in Samsun tobacco sap, stained with phosphotungstate. Bar represents 300 nm.
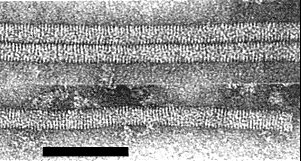
Particles of a ribgrass mosaic isolate from Scotland, stained with uranyl formate. Bar represents 100 nm. (Courtesy of I. M. Roberts.)

Rounded plates (P) in hair cells of Nicotiana tabacum in face view (left) and in side view (right). (Courtesy of D. Milicic.)

Polyacrylamide gel electrophoretic patterns of the virus particle protein of the following isolates: wasabi (W), TMV-C (C), type ribgrass mosaic (HR), lychnis (Ly), and an ordinary strain of TMV (OM). (Courtesy of Y. Ohashi.)
References list for DPV: Ribgrass mosaic virus (152)
- Chessin, Zaitlin & Solberg, Phytopathology 57: 452, 1967.
- Goldin, C. r. Acad. Sci. U.R.S.S., N.S. 88: 933, 1953.
- Goto & Oshima, Ann. phytopath. Soc. Japan 27: 109, 1962.
- Hollings, Acta Hort. 36: 23, 1974.
- Holmes, Phytopathology 31: 1089, 1941.
- Holmes, Phytopathology 40: 487, 1950.
- Juretic, Phytopath. Z. 79: 16, 1974.
- Juretic, Milicic & Mamula, Acta bot. croat. 32: 29, 1973.
- Juretic & Wetter, Proc. 7th Conf. Czechoslov. Pl. Virol, High Tatras, 1971: 369, 1973.
- Juretic, Wrischer & Polák, Biológia Pl. 11: 284, 1969.
- Knight, J. biol. Chem. 197: 241, 1952.
- Knight & Lauffer, J. biol. Chem. 144: 411, 1942.
- Kovachevsky, Phytopath. Z. 49: 127, 1963.
- Kovachevsky, Proc. 6th Conf. Czechoslov. Pl. Viral., Olomouc, 1967: 250, 1969.
- Milicic, Stefanac, Juretic & Wrischer, Virology 35: 356, 1968.
- Oshima, Ohashi & Umekawa, Ann. phytopath. Soc. Japan 37: 319, 1971.
- Oshima, Ohashi & Umekawa, Ann. phytopath. Soc. Japan 40: 243, 1974.
- Oster, J. biol. Chem. 190: 55, 1951.
- Pei, Acta microbiol. sin. 8: 420, 1962.
- Polák, Proc. 5th Conf. Czechoslov. Pl. Viral., Prague, 1962: 168, 1962.
- Schumann, Phytopath. Z. 48: 1, 1963.
- Siegel & Wildman, Phytopathology 44: 277, 1954.
- Tochihara, Komuro & Kobari, Proc. Kanto Pl. Prot. Soc. 11: 46, 1964.
- Valleau & Johnson, Phytopathology 33: 210, 1943.
- van Regenmortel, Virology 31: 467, 1967.
- Wittmann, Hindennach & Wittmann-Liebold, Z. Naturforsch. B24: 877, 1969.