Details of DPV and References
DPV NO: 154 October 1975
Family: Virgaviridae
Genus: Tobamovirus
Species: Cucumber green mottle mosaic virus | Acronym: CGMMV
Cucumber green mottle mosaic virus
M. Hollings Glasshouse Crops Research Institute, Littlehampton, Sussex BNI6 3PU, England
Y. Komuro Institute for Plant Virus Research, 959 Aobacho, Chiba 280, Japan
H. Tochihara Institute for Plant Virus Research, 959 Aobacho, Chiba 280, Japan
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
Described by
Ainsworth (1935).
Synonyms
- Cucumber virus 3
(Ainsworth, 1935)
- Cucumis virus 2 (Smith, 1957)
-
An RNA-containing virus found in cucurbitaceous crops in Europe, India and Japan. The infective particles are tubules c. 300 x 18 nm. The virus is readily sap-transmissible, but most strains have a restricted host range. Transmitted by foliage contact, handling of plants during cultivation, soil contamination, or through cucurbit seeds; no biological vector is known.
Main Diseases
In cucumber (Cucumis sativus) the type strain causes leaf mottling, blistering and distortion (Fig. 4), with stunted growth. Yield losses may be 15% (Fletcher et al., 1969); fruits are usually unmarked (Smith, 1957), but some strains cause severe fruit mottling and distortion (Fig. 1) (Inouye et al., 1967a). Some Asian cvs show no leaf symptoms but suffer yield losses (Kooistra, 1968).
The aucuba mosaic strain produces bright yellow leaf mottling, with only slight leaf distortion and stunting; fruits may show yellow or silver-coloured streaks and flecks, especially at higher temperatures (Smith, 1957).
In watermelon (Citrullus vulgaris), the watermelon strain produces slight leaf mottling and dwarfing, but infection at fruit-set, or shortly afterwards, can induce serious internal discoloration and decomposition in the fruit (Komuro et al., 1971).
Geographical Distribution
Europe, India and Japan.
Host Range and Symptomatology
Found in naturally infected cucumber, watermelon and melon (Cucumis melo) crops, but apparently not in vegetable marrow (Cucurbita pepo). Occurs also in bottlegourd (Lagenaria siceraria) used as rootstocks for watermelon cultivation.
Most UK isolates tested are apparently restricted in host range to Cucurbitaceae (M.
Hollings, unpublished data); some strains from Japan and India and one isolate from UK
induce local lesions in Chenopodium amaranticolor and/or Datura stramonium
(Komuro et al., 1971;
Tochihara & Komuro, 1974).
East European isolates
of the aucuba mosaic strain are reported to give chlorotic local lesions in tobacco cvs
Samsun and Xanthi-nc
(Brcák et al., 1962),
and systemic mottling in
Chenopodium murale.
Diagnostic species
Propagation species
Assay species
Strains
Several strains are distinguished serologically and by differential responses in C. amaranticolor and D. stramonium (Tochihara & Komuro, 1974). The principal strains are:-
Type strain (Ainsworth, 1935) = cucumber green mottle mosaic virus, reported in UK and Europe. Does not normally cause fruit symptoms in cucumber, and produces a few local lesions in Chenopodium amaranticolor under certain conditions only. No infection in Datura stramonium and Petunia hybrida.
Cucumber aucuba mosaic strain (Bewley, 1923) = cucumber virus 4 (Ainsworth, 1935) = Cucumis virus 2A (Smith, 1957). Reported in UK and Europe; similar isolates noted in India (Vasudeva et al., 1949). Can induce conspicuous fruit symptoms in cucumber, and causes local lesions in Chenopodium amaranticolor but not in Datura stramonium.
Watermelon strain: recorded in Japan (Komuro et al., 1971). Induces local lesions in C. amaranticolor (Fig. 2) but not in D. stramonium.
Japanese cucumber strain: recorded in Japan (Komuro et al., 1971). Causes severe fruit distortion in cucumber and induces local lesions in D. stramonium (Fig. 3) but not in C. amaranticolor.
Yodo strain: recorded from cucumbers at Yodo, Japan (Kitani et al., 1970; Tochihara & Komuro, 1974). Causes fruit distortion in cucumber, and local lesions in C. amaranticolor, D. stramonium and Petunia hybrida.
Indian strain C (=2C): from bottlegourd in India (Vasudeva et al., 1949), causes blister-mottle, stunting and loss of crop; induces local lesions in C. amaranticolor, symptomless infection of inoculated leaves of D. stramonium and no infection in tobacco or Petunia hybrida.
Transmission by Vectors
No confirmed reports. No transmission was obtained with the aphids Myzus persicae and Aphis gossypii (Inouye et al., 1967a), or with cucumber leaf beetles (Aulacophora femoralis) (Nagai et al., 1974). Infection occurs via the roots in soil contaminated by infective plant debris (Inouye et al., 1967b; Nagai et al., 1974), but may be prevented by soil fumigation with methyl bromide (Inouye et al., 1967c). Transmission also occurs through foliage contact, when plants are handled during cultivation, or when infected rootstocks are used in watermelon or cucumber cultivation.
Transmission through Seed
In cucumber, up to 8% seed transmission occurs up to 1 month after harvesting seed, falling to c. 1% after 5 months storage of seed (van Koot & van Dorst, 1959). Seed transmission also occurs in bottlegourd, and up to 5% in watermelon, although the virus was not detected by inoculation from watermelon seeds (Komuro et al., 1971; Nagai et al., 1974). Contamination of seeds is mostly external, and can be eliminated by dry heat-treatment of seeds for 3 days at 70°C (Fletcher et al., 1969; Nagai et al., 1974) without impairing seed germination. Pollen contains a low concentration of the virus.
Transmission by Dodder
Cucumber green mottle mosaic virus is transmitted by Cuscuta subinclusa, C. lupuliformis and C. campestris (Schmelzer, 1956), and the aucuba mosaic strain by C. campestris (Johnson, 1941).
Serology
The virus is a good immunogen; antisera prepared by one intravenous plus two intramuscular injections (with complete adjuvant) had titres in precipitin tube tests of 1/4096, the precipitates being flocculent (flagellar) (M. Hollings, unpublished data). Also reacts well in microprecipitin tests, and in immunodiffusion using freshly prepared 0.8% agar gel; it produces one major, and occasionally one minor, line of precipitation. Purified virus preparations mixed with an equal volume of glycerol and stored at c. 2°C retain nearly all serological activity for at least 6 years (M. Hollings, unpublished data).
Relationships
Infective particles of the different strains of this virus appear morphologically indistinguishable from those of other tobamoviruses, and are serologically related to several of them. Cross-protection tests with other tobamoviruses have given conflicting results, but usually slight or no evident protection has been found between cucumber and typical tobacco (vulgare) types (Fulton, 1950; Knight, 1955; Rochow, 1956; Brcák et al., 1962; Wu & Wildman, 1963).
The watermelon strain is serologically fairly close to two cucumber strains from UK and to strain C from India; these are much more distantly related to the Japanese cucumber and Yodo strains and to tobacco mosaic virus (type strain), and very distantly related to tomato mosaic virus and other tobamoviruses (Fig. 5) (Nozu et al., 1971; Tochihara & Komuro, 1974; van Regenmortel, 1975; M. Hollings, unpublished data). The Japanese cucumber and Yodo strains are also closely related to each other.
Stability in Sap
All strains are extremely stable: in Cucumis sativus sap, the type strain loses infectivity after 10 min at 90°C (Smith, 1957), watermelon and Yodo strains at 90-100°C (Kitani et al., 1970; Nagai et al., 1974), and the Indian strain C at 86-88°C (Vasudeva et al., 1949).
The dilution end-point ranges from 10-6 for type strain to 10-7 for the watermelon strain (Nagai et al., 1974). Infectivity survives for several months at laboratory temperatures, and several years at 0°C.
Purification
The virus is easily purified from systemically infected cucumber plants. Methods include:-
1. (Tung & Knight, 1972). Harvest plants c. 3 weeks after infection, blend in 0.1 M phosphate buffer pH 7.0 (2 ml/g tissue) and subject to 2-3 cycles of differential centrifugation. Adsorbed pigments are then removed by homogenizing the virus preparation in 0.1 M phosphate buffer pH 7.0 + equal volume of a 1:1 (v/v) mixture of n-butanol and chloroform for 2-3 min at laboratory temperature. The aqueous layer containing the virus is separated by centrifugation at 900 g and given two more cycles of differential centrifugation. The final pellet is re-suspended in distilled water.
2. Pure preparations can be obtained without the use of an ultracentrifuge (M. Hollings & R. Barton, unpublished data). Harvest plants 3-4 weeks after infection, freeze leaves overnight, then blend with 0.05 M phosphate buffer pH 7.6 (4 ml buffer/g tissue) containing 0.1% (v/v) thioglycollic acid. Filter through cloth. Add n-butanol dropwise (9.3 ml/100 ml juice) and shake the mixture for 45 min at laboratory temperature. Centrifuge at 10,000 g for 30 min. Precipitate the virus from the supernatant fluid by adding 2% (w/v) NaCl then 4% (w/v) polyethylene glycol (M. Wt 6000), stand the mixture ½-1 h at c. 4°C, and centrifuge for 10 min at 10,000 g. Re-suspend the precipitate in distilled water (1 ml/25 g original tissue) and remove insoluble material (5 min at 10,000 g). Apply 1-1.5 ml of this preparation to a column (85 x 1.5 cm) of controlled-pore glass beads (70 nm pore size; Sigma London Chemical Co. Ltd.), and elute with 0.04 M phosphate buffer (pH 7.0). The virus elutes in the void volume, well separated from host material, and can be further concentrated by precipitation with 2.5 vol ethanol.
Properties of Particles
Purified preparations sediment as a major infective component (Fig. 7), with minor components (probably dimers and trimers). Sedimentation coefficient (s°20,w): 185 S (M. Hollings & A. A. Brunt, unpublished data); 195 S (Brcák et al., 1962).
Isoelectric point: about pH 4.98 (Oster, 1951; Brcák et al., 1962).
A260/A280: 1.38. Amax(260)/Amin(249): 1.05. (Values corrected for light-scattering; M. Hollings, unpublished data).
Particle Structure
Infective particles are straight rods (Fig. 6), c. 300 x 18 nm when mounted in 2% phosphotungstate, with helical structure of pitch 23 Å and a central canal of radius 20 Å. The RNA lies at c. 40 Å radius from the centre of the core, and the helix comprises 49 subunits/3 turns (Holmes & Franklin, 1958).
Particle Composition
Nucleic acid: RNA, single-stranded, with nucleotide base ratios G25; A26; C19; U30 moles % (Linnasalmi & Toiviainen, 1974). Comprises c. 6% of particle weight (Knight, 1954).
Protein: comprises c. 94% of particle weight. Three isolates from cucumber each contained 15 amino acids, with an average of 160 residues/subunit; M. Wt c. 17,200 (Linnasalmi & Toiviainen, 1974). The amino acid compositions of these three European isolates from cucumber were very similar to those of the watermelon strain (Nozu et al., 1971), and strain CV-4 (aucuba mosaic) (van Regenmortel, 1967). Van Regenmortel's isolate, however, is regarded as a typical green mottle mosaic strain by Tung & Knight (1972) and by Linnasalmi & Toiviainen (1974). An authentic aucuba mosaic strain shows some differences from these (Tung & Knight, 1972).
Partial amino acid sequences for the proteins of the Japanese cucumber and watermelon strains are given by Funatsu (1967) and Nozu et al. (1971). Substantial differences (11 substitutions) were recorded between the sequences of amino acids 91 to 122.
Relations with Cells and Tissues
The virus is present in all tissues, including pollen, and occasionally in seed embryos. Cytological abnormalities, including mitochondrial vesiculation, have been reported in association with the cucumber green mottle mosaic strain (Hatta et al., 1971; Hatta & Ushiyama, 1973).
There are conflicting reports regarding intracellular inclusions: Bawden & Sheffield (1939) found none in cucumbers containing green mottle mosaic or aucuba mosaic strains, but Brcák & Hrsel (1961) found cytoplasmic inclusions similar to those induced by type strain of tobacco mosaic virus in cucumber cells containing the aucuba mosaic strain; Milicic (1969) recorded circular plate inclusions with the green mottle mosaic strain.
Notes
It is seldom possible to identify this virus with certainty from symptoms in cucurbitaceous plants or by indicator plant reactions, for similar symptoms can be caused by other viruses. Cucumber mosaic and alfalfa mosaic viruses, however, readily infect many solanaceous test plants systemically; these and watermelon mosaic virus are transmitted non-persistently by aphids. Some strains of watermelon mosaic virus have a restricted host-range similar to that of the type strain of cucumber green mottle mosaic virus. Squash mosaic virus is beetle-transmitted and is restricted to Cucurbitaceae.
Serological tests offer positive identification of strains of cucumber green mottle mosaic virus, which have particles unlike any of the above four viruses.
Acknowledgements
Photographs 1, 2, 3 and 5, Institute for Plant Virus Research; 4, 6 and 7, Glasshouse Crops Research Institute.
Figures

Distorted cucumber fruit infected with Japanese cucumber strain.

Local lesions caused in Chenopodium amaranticolor by the watermelon strain.

Chlorotic local lesions caused in Datura stramonium by the Japanese cucumber strain.

Systemic light and dark green blister mottle and malformation in leaf of cucumber (cv. Butcher’s Disease Resister) infected with typical UK strain (right); healthy leaf (left).

Immunodiffusion plate, illustrating relationships among strains of the virus. Central well - antiserum to watermelon strain. Antigens in the peripheral wells: C = Indian strain C; Wa = watermelon strain; T = tomato mosaic virus; OM = ordinary tobacco TMV; Y = Yodo strain; Cu - cucumber strain (Japanese); E1 and E2 = cucumber strains (UK).
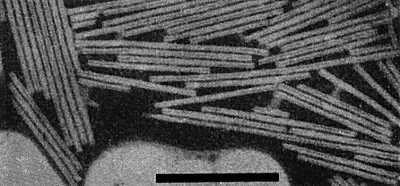
Virus particles mounted in 2% potassium phosphotungstate. Bar represents 300 nm.

Sedimentation pattern of purified virus preparation in the analytical ultracentrifuge. Sedimentation is from left to right.
References list for DPV: Cucumber green mottle mosaic virus (154)
- Ainsworth, Ann. appl. Biol. 22: 55, 1935.
- Bawden & Sheffield, Ann. appl. Biol. 26: 102, 1939.
- Bewley, Diseases of Glasshouse Plants, Benn Bros., London, 1923.
- Brcák & Hrsel, Biológia Pl. 3: 132, 1961.
- Brcák, Ulrychová & Cech, Virology 16: 105, 1962.
- Fletcher, George & Green, Pl. Path. 18: 16, 1969.
- Fulton, Phytopathology 40: 219, 1950.
- Funatsu, J. agric. Chem. Soc. Japan 41: 53, 1967.
- Hatta & Ushiyama, J. gen. Virol. 21: 9, 1973.
- Hatta, Nakamoto, Takagi & Ushiyama, Virology 45: 292, 1971.
- Hollings, Acta Hort. 36: 23, 1974.
- Holmes & Franklin, Virology 6: 328, 1958.
- Inouye, Inouye, Asatani & Mitsuhata, Nogaku Kenkyu 51: 175, 1967a.
- Inouye, Inouye, Asatani & Mitsuhata, Nogaku Kenkyu 51: 187, 1967b.
- Inouye, Inouye, Asatani & Mitsuhata, Nogaku Kenkyu 51: 199, 1967c.
- Johnson, Phytopathology 31: 649, 1941.
- Kitani, Kiso & Shigematsu, Proc. Assoc. Pl. Prot. Sikoku 5: 59, 1970.
- Knight, Adv. Virus Res. 2: 153, 1954.
- Knight, Virology 1: 261, 1955.
- Komuro, Tochihara, Fukatsu, Nagai & Yoneyama, Ann. phytopath. Soc.Japan 37: 34, 1971.
- Kooistra, Euphytica 17: 136, 1968.
- Linnasalmi & Toiviainen, Ann. Agric. Fenn. (Phytopath.) 13: 79, 1974.
- Milicic Zast. Bilja 20: 101, 1969.
- Nagai, Toki & Fukatsu Bull. Chiba-ken agric. Exp. Stn 15: 1, 1974.
- Nozu, Tochihara, Komuro & Okada, Virology 45: 577, 1971.
- Oster, J. biol. Chem. 190: 55, 1951.
- Rochow, Phytopathology 46: 133, 1956.
- Schmelzer, Phytopath. Z. 28: 1, 1956.
- Smith, A Textbook of Plant Virus Diseases, 2nd ed., Churchill. 1957.
- Tochihara & Komuro, Ann. phytopath. Soc. Japan 40: 52, 1974.
- Tung & Knight, Virology 48: 574, 1972.
- van Koot & van Dorst, Tijd. PlZkt. 65: 257, 1959.
- van Regenmortel, Virology 31: 467, 1967.
- van Regenmortel, Virology 64: 415, 1975.
- Vasudeva, Raychaudhuri & Singh, Indian Phytopath. 2: 180, 1949.
- Wu & Wildman, Nature, Lond. 199: 1015, 1963.