Details of DPV and References
DPV NO: 157 September 1976
Family: Closteroviridae
Genus: Closterovirus
Species: Wheat yellow leaf virus | Acronym: WYLV
Wheat yellow leaf virus
T. Inouye College of Agriculture, University of Osaka Prefecture, Sakai, Osaka, Japan
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
Described by
Inouye et al. (1973).
A virus with very flexuous elongated particles c. 1600-1850 nm long. Transmitted by aphids in a semi-persistent manner but not by inoculation of sap. Infects only species of Gramineae, and occurs in Japan. A typical, distinct member of the closterovirus group.
Main Diseases
In wheat, barley and rye, the virus causes diffuse chlorotic flecks or interveinal chlorosis spreading from the leaf tips, then severe yellowing of the entire leaves, sheaths and heads, usually followed by rapid death of infected tissues (Fig. 1, Fig. 3, Fig. 4). Affected immature heads often ripen prematurely, and there is appreciable loss of yield. The virus causes mild yellowing in Agropyron kamoji and red leaf symptoms in Italian ryegrass. Infected Agropyron kamoji may be an important reservoir host of the virus.
Geographical Distribution
Japan (Inouye et al., 1973).
Host Range and Symptomatology
The virus infects several species within the Gramineae. Susceptible hosts known are Agropyron kamoji, Avena sativa, Hordeum vulgare, Lolium multiflorum, Secale cereale and Triticum aestivum (Inouye et al., 1973).
-
Diagnostic species
- Triticum aestivum
(wheat). Diffuse chlorotic flecks (Fig. 2), then pronounced yellowing of leaves spreading from the tips, about 10-15 days after inoculation by aphids, followed by rapid death of affected leaves. Leaves produced later on show symptoms only at the tips. Young seedlings may be killed or severely stunted, producing only a few tillers. - Avena sativa (oat). Orange to red discoloration of leaves, about 10-15 days after inoculation by aphids, followed by death of affected leaves. Leaves produced later on show symptoms only at the tips.
-
Propagation species
- Triticum aestivum
is a good source of virus for propagation. For maintaining cultures, Agropyron kamoji is a suitable host.Assay species
- No known local lesion host. Triticum aestivum and Avena sativa are good for insect transmission tests.
Strains
No strains reported.
Transmission by Vectors
Transmitted by the aphid Rhopalosiphum maidis in a semi-persistent manner (Inouye et al., 1973). Vector aphids can transmit the virus after acquisition feeds longer than 3 h and test feeds longer than 1 h, and can retain the virus for up to 2 days. Efficiency of transmission by vector aphids increases with increase of acquisition feed beyond 4 h and of test feed beyond 2 h.
Transmission through Seed
No seed transmission was found in 638 seedlings of wheat (Inouye et al., 1973).
Serology
No antisera have been prepared.
Relationships
The virus is distantly related serologically to carnation yellow fleck virus (Smookler & Loebenstein, 1974), a virus very similar serologically to carnation necrotic fleck virus (Inouye, 1974; T. Inouye et al., unpublished data). It resembles festuca necrosis virus (Schmidt et al., 1963) in having very flexuous particles about 1.7 µm long, in its transmissibility by aphids, and in producing red leaf symptoms in oat; however, festuca necrosis virus is reported not to infect barley. The virus also resembles beet yellows (Russell, 1970), beet yellow stunt (Duffus, 1972; Hoefert et al., 1970), carnation necrotic fleck (Inouye, 1974), and citrus tristeza (Price, 1970) viruses in its particle morphology, in its transmission by aphids in the semi-persistent manner, and in its relations with tissues. All belong to the closterovirus group.
Stability in Sap
Not tested.
Purification
The virus is partially purified by the following procedure (T. Inouye et al., unpublished data). Grind frozen leaves in 0.1 M Tris-citrate buffer (3 ml/g), adjusted to pH 8.2 using propanediamine. Add 2 M CaCl2 (1 ml/50 ml of extract) and 2 M K2HPO4 (2 ml/50 ml of extract) to the crude extract and clarify by low speed centrifugation. Precipitate the virus by adding Triton X-100 to 0.5% and polyethylene glycol (PEG M. Wt 6000) to 6%, and resuspend in 0.01 M buffer containing 0.1% Triton X-100 and 0.1% 2-mercaptoethanol. Purify further by centrifugation through a 30% sucrose cushion followed by sucrose density gradient centrifugation.
Properties of Particles
Unknown.
Particle Structure
Particles are very flexuous filaments c. 1600-1850 nm long and c. 10 nm in diameter (Fig. 5), and are constructed helically with the pitch of the basic helix c. 3.4 nm (Inouye et al., 1973).
Particle Composition
Unknown.
Relations with Cells and Tissues
Virus particles are found only in the phloem sieve tubes and phloem parenchyma. Some phloem cells are almost filled with the masses of filamentous particles (Fig. 6). Causes necrosis of some phloem cells (Inouye et al., 1973).
Notes
In Japan, wheat yellow leaf virus often occurs together with barley yellow dwarf virus (Rochow, 1970), from which it can be freed by allowing vector aphids an acquisition feed of about 4 h followed immediately by a test feed of about 4 h. The virus can also be distinguished from barley yellow dwarf virus by particle morphology and symptomatology; it causes pronounced yellowing of leaves followed by rapid death of the entire leaves and stems in wheat, whereas the yellowing caused by barley yellow dwarf virus is often restricted to the tips and margins of the leaves. Wheat yellow leaf virus can be easily distinguished from the other viruses infecting wheat and barley in Japan, such as barley yellow mosaic, barley stripe mosaic, soil-borne wheat mosaic and wheat yellow mosaic viruses by symptomatology, particle morphology and aphid transmissibility.
Figures

Leaves of naturally infected wheat; (left) two leaves showing interveinal chlorosis; (centre) two leaves showing pronounced chlorotic flecks; (right) three leaves showing severe yellowing.

Leaves of wheat plants infected by aphid inoculation showing chlorotic flecks spreading from the leaf tips.

Naturally infected wheat showing chlorotic flecks and yellowing of flag leaf and sheath; (right) healthy, others infected.

Naturally infected barley; (right) healthy, others infected.
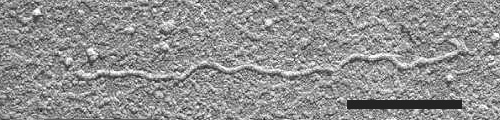
Shadowcast virus particle in dip preparation. Bar represents 500 nm.

Aggregates of virus particles in a portion of a sieve tube of wheat. Bar represents 500 nm.
References list for DPV: Wheat yellow leaf virus (157)
- Duffus, Phytopathology 62: 161, 1972.
- Hoefert, Esau & Duffus, Virology 42: 814, 1970.
- Inouye, CMI/AAB Descriptions of Plant Viruses 136 , 3 pp., 1974.
- Inouye, Mitsuhata, Heta & Hiura, Nogaku Kenkyu 55: 1, 1973.
- Price, CMI/AAB Descriptions of Plant Viruses 33, 3 pp., 1970.
- Rochow, CMI/AAB Descriptions of Plant Viruses 36, 4 pp., 1970.
- Russell, CMI/AAB Descriptions of Plant Viruses 13, 3 pp., 1970.
- Schmidt, Richter, Hertzsch & Klinkowski, Phytopath. Z. 47: 66, 1963.
- Smookler & Loebenstein, Phytopathology 64: 979, 1974.