Details of DPV and References
DPV NO: 165 September 1976
Family: Unallocated ssRNA+ viruses
Genus: Idaeovirus
Species: Raspberry bushy dwarf virus | Acronym: RBDV
There is a more recent description of this virus: DPV 363
Raspberry bushy dwarf virus
A. F. Murant Scottish Horticultural Research Institute, Invergowrie, Dundee, Scotland
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
Described by
Cadman (1961) and
Barnett & Murant (1970).
Synonym
- Loganberry degeneration virus (Rev. appl. Mycol. 39: 603)
-
An RNA-containing virus with quasi-isometric particles about 33 nm in diameter, widespread throughout the world in some cultivars of red and black raspberry. It is readily transmissible by inoculation of sap and infects a wide range of herbaceous plants, often causing no symptoms. The virus is seed-borne and pollen-borne in raspberry, and spreads from plant to plant by pollination. No vector is known.
Main Diseases
The virus is one of several associated with ‘bushy dwarf’ (Cadman & Harris, 1951) or ‘symptom-less decline’ (Cadman, 1952) of Lloyd George raspberry (Rubus idaeus); affected plants produce few, stunted canes and are very prone to autumn fruiting. However, raspberry bushy dwarf virus does not, despite its name, cause this disease (Barnett & Murant, 1970) although it may be a contributory factor when occurring together with aphid-borne latent viruses such as black raspberry necrosis virus (‘52 V’; Jones, 1976). Fruit on infected plants may have a high proportion of aborted drupelets (Murant, Chambers & Jones, 1974). The virus occurs symptomlessly in many other R. idaeus cultivars (including Canby, Carnival, Creston, Newburgh and Norfolk Giant), in loganberry and boysenberry, and in the black raspberry (R. occidentalis) cvs. Munger, New Logan and Plum Farmer (Barnett & Murant, 1970; Converse, 1973; Murant et al., 1974). Following graft inoculation the virus induces transient mild chlorotic mottle, watermarks or line-patterns in R. laciniatus, R. procerus and R. phoenicolasius (Jones, 1976).
Geographical Distribution
Found in western Europe, North America and New Zealand. Probably occurs wherever susceptible Rubus cultivars are grown.
Host Range and Symptomatology
The virus infected 55 species in 12 dicotyledonous families, most of them symptomlessly (Barnett & Murant, 1970, 1971). It causes mild systemic mottle symptoms in some species of Chenopodiaceae and local lesions in some species of Leguminosae. The virus is readily transmissible between herbaceous plants by inoculation of sap. Transmission from Rubus is easier in spring and autumn than in summer and is best achieved by grinding the leaves in 2% nicotine solution in phosphate buffer, pH 7.
-
Diagnostic species
- Chenopodium amaranticolor
. Transient chlorotic local lesions may form in 4 days especially in spring and autumn; systemic chlorotic rings and line patterns develop after 7 days (Fig. 1). -
C. murale. Local sunken necrotic rings
(Fig. 3).
No systemic infection.
- C. quinoa. Transient chlorotic local lesions may form in 4-7 days especially in spring and autumn; systemic chlorotic spots develop after 7 days followed by light and dark green mosaic and ring patterns (Fig. 2).
- Phaseolus vulgaris (French bean) cv. The Prince. In winter, small brown local lesions develop after 3 days (Fig. 4). No systemic infection.
- Nicotiana clevelandii. Symptomless systemic infection.
- C. quinoa. Transient chlorotic local lesions may form in 4-7 days especially in spring and autumn; systemic chlorotic spots develop after 7 days followed by light and dark green mosaic and ring patterns (Fig. 2).
-
Propagation species
- Chenopodium quinoa
is suitable for maintaining cultures and as a source of virus for purification.Assay species
- Phaseolus vulgaris
cv. The Prince is useful in winter or when grown in controlled environment chambers (20°C, 5000 lux, 16 h photoperiod) but lesion development is inhibited by a component of C. quinoa sap which occurs in increased concentrations in plants grown in long days and high light intensities (Barnett & Murant, 1970). Lesion numbers are increased by keeping beans in the dark for 1 day before inoculation and by using phosphate buffer in the inoculum. Chenopodium murale is more reliable but the lesions are less easy to count.
Strains
All tested isolates from Rubus idaeus are serologically identical but two isolates from R. occidentalis differed slightly from R. idaeus isolates in serological tests and in properties in vitro (Jones & Murant, 1972a; Murant & Jones, 1976). Isolates differ in virulence and tend to become ‘attenuated’ after prolonged culture in Chenopodium quinoa.
Transmission by Vectors
No vector is known. None of the insects commonly present on raspberry in Scotland transmitted the virus (Cadman, 1970).
Transmission through Seed
Up to 77% of seedlings from infected R. idaeus plants may be infected (Cadman, 1965; Converse, 1973; Murant et al., 1974); the virus is transmitted through both gametes. Up to 15% seed transmission was found in R. phoenicolasius (A. T. Jones, unpublished data). Only 1-2% seed transmission was detected in Fragaria vesca, and none in Chenopodium quinoa (Murant et al., 1974).
In raspberry, the virus carried in pollen infects not only the progeny seedlings but also the pollinated plant (Cadman, 1965; Murant et al., 1974). This seems to be the only method of spread in the field because, in plots containing infector plants, no spread was detected to healthy Lloyd George raspberry plants that were prevented from flowering for 3 years; in contrast, most healthy plants that were allowed to flower became infected during the first two flowering seasons (Murant et al., 1974).
Healthy or infected raspberry flowers pollinated with infected pollen may produce ‘crumbly’ fruit containing a high proportion of aborted drupelets (Murant et al., 1974).
Transmission by Dodder
No information.
Serology
Moderately immunogenic in rabbits; antiserum with a titre of 1/512 was produced in a rabbit following four intravenous injections of 2.5 mg virus at 2-3 day intervals. Microprecipitin tests or double diffusion tests in 0.5% agarose gel are satisfactory. In immunodiffusion tests the virus gives a single line with moderate dilutions of antiserum but some antisera may give additional lines at low dilutions suggesting that smaller virus-related components exist.
Relationships
Despite earlier reports (Cadman, 1963; 1970) raspberry bushy dwarf virus is unrelated to the filamentous virus apple chlorotic leaf spot, which has never been found in Rubus. Raspberry bushy dwarf virus also failed to react with antisera to twenty-four isometric viruses including seven ilarviruses (Barnett & Murant, 1970). Its particle morphology and mode of transmission through pollination suggest affinities with the ilarvirus group but it differs from these viruses in sedimentation behaviour and RNA composition.
Stability in Sap
In Chenopodium quinoa sap, a typical Rubus idaeus isolate lost infectivity when diluted 10-4, heated for 10 min at 65°C or stored for 4 days at 22°C. Infectivity was stabilised by 2-mercaptoethanol but not by sodium diethyl dithiocarbamate (Barnett & Murant, 1970). An isolate from R. occidentalis had a dilution end-point of only 10-2 in C. quinoa sap and lost infectivity after 2-3 h at room temperature (Jones & Murant, 1972a; Murant & Jones, 1976).
Purification
The following modification (A. F. Murant, unpublished data) of the procedure of Barnett & Murant (1970) is now used: Harvest inoculated and systemically infected Chenopodium quinoa leaves about 10 days after inoculation and homogenize each 100 g tissue in 400 ml 0.05 M sodium phosphate buffer, pH 7.0, containing 0.02 M 2-mercaptoethanol. Adjust the pH to 4.8 with 0.5 M citric acid and centrifuge 20 min at 3000 g. In some preparations the virus is in the supernatant fluid after this step but in others it is found in the pellet. If the virus is in the supernatant fluid, precipitate it by adding polyethylene glycol (PEG), M. Wt 6000, to 8% (w/v) and NaCl to 0.8% (w/v) and centrifuging at low speed. Use 0.05 M sodium citrate buffer, pH 6.0, containing 0.02 M 2-mercaptoethanol (2-ME) to resuspend the pellets and throughout all subsequent stages. If the virus is precipitated by treatment at pH 4.8 it may be recovered by extracting the pellets overnight in citrate buffer + 2-ME and removing insoluble material by low speed centrifugation. It is then treated with polythylene glycol, as above. Concentrate the virus by two or more cycles of differential centrifugation. Further purification is achieved by exclusion chromatography in columns (80 x 1.6 cm) of 2% agarose beads and by sucrose density gradient centrifugation. Yields may be up to 5 mg virus per 100 g leaf material but are usually much lower (0.5-1 mg).
Properties of Particles
The particles sediment to form a rather broad zone in sucrose density gradients. They are unstable and readily disrupt in many commonly used buffer systems, and in the presence of as little as 0.01% sodium dodecyl sulphate, suggesting that they are stabilized by protein-RNA linkages (A. F. Murant, unpublished data).
Sedimentation coefficient, s°20,w: 115 S in 0.05 M citrate buffer at pH 6 or 7. A minor component of 111 S is detectable in some preparations. Occasionally, non-infective particles sedimenting at 34 S are found.
Formaldehyde-fixed virus centrifuged to equilibrium at 20°C in solutions of CsCl or RbBr forms a band of aggregated particles at a position corresponding to a density of 1.37 g/ml (Murant, 1975).
A260/A280: 1.62 (Barnett & Murant, 1970).
In immunoelectrophoresis in 0.7% agarose in 0.025 ionic strength phosphate buffer pH 6, 7 or 8 the virus migrated towards the anode as a single component (A. F. Murant, unpublished data).
Particle Structure
The particles are quasi-isometric c. 33 nm in diameter (Fig. 6). They disrupt in phosphotungstate but are preserved in uranyl acetate or uranyl formate (Barnett & Murant, 1970). Occasionally preparations contain particles c. 17 nm in diameter (the 34 S particles) which are serologically identical to the 33 nm particles but are not infective.
Particle Composition
Nucleic acid: RNA, single-stranded, about 24% of particle weight estimated from the buoyant density in CsCl. Polyacrylamide gel electrophoresis of undenatured RNA (Fig. 5) reveals three components of estimated M. Wt 2.0, 0.8 and 0.3 x 106 (Murant, 1975 and unpublished data).
Protein: Particles contain a major polypeptide whose relative mobility in polyacrylamide/sodium dodecyl sulphate gels depends on gel concentration; its estimated M. Wt after allowing for this effect is 29,000 (Murant, 1975). Minor amounts of smaller polypeptides occur even in highly purified preparations but their significance is not known.
Relations with Cells and Tissues
No information on cytopathological effects. The virus was eliminated from R. idaeus (Murant et al., 1974; F. C. Mellor & R. Stace-Smith, personal communication) and from R. occidentalis (Converse, 1973) by heat therapy.
Notes
The characteristic symptoms in Chenopodium quinoa and lack of symptoms in Nicotiana clevelandii distinguish raspberry bushy dwarf virus from most nepoviruses found in Rubus, except perhaps strawberry latent ringspot virus; the nepoviruses also differ from it in particle morphology (regular isometric particles with hexagonal outlines, some penetrated by negative stain). The type of symptoms in C. quinoa also distinguishes it from black raspberry necrosis virus (‘52V’ virus of Jones & Murant, 1972b), which is difficult to transmit by manual inoculation and has regular isometric particles present in very low concentrations in sap. Black raspberry latent virus (Lister & Converse, 1972) resembles raspberry bushy dwarf virus in particle morphology and transmission during pollination but causes severe systemic necrosis in C. quinoa and Phaseolus vulgaris.
Because of its mode of transmission, raspberry bushy dwarf virus can at present only be controlled by use of immune cultivars. Although the virus causes only mild leaf symptoms, its effects on fruit quality and its possible interaction with other viruses point to the undesirability of introducing cultivars that are susceptible.
Figures

Systemic chlorotic rings and line patterns in Chenopodium amaranticolor.

Systemic chlorotic rings and line patterns in C. quinoa.

Local lesions in C. murale.
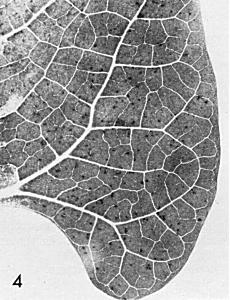
Local lesions in Phaseolus vulgaris cv. The Prince.
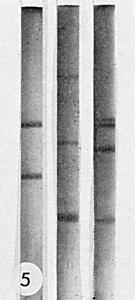
Electrophoresis of RNA in 2.4% polyacrylamide gels in 0.036 M Tris, 0.03 M Na2HPO4, 0.001 M disodium ethylenediamine-tetraacetate, pH 7.8. A current of 5 mA/gel was applied for 2 h. The direction of migration was from top to bottom, (Left), Escherichia coli RNA; (Centre), RNA of raspberry bushy dwarf virus, (Right), RNA of brome mosaic virus.

Particles of raspberry bushy dwarf virus in uranyl formate. Bar represents 100 nm.
References list for DPV: Raspberry bushy dwarf virus (165)
- Barnett & Murant, Ann. appl. Biol. 65: 435, 1970.
- Barnett & Murant, Annls Phytopath. Numero hors série: 129, 1971.
- Cadman, Ann. appl. Biol. 39: 501, 1952.
- Cadman, Hort. Res. 1: 47, 1961.
- Cadman, Pl. Dis. Reptr 47: 459, 1963.
- Cadman, Pl. Dis. Reptr 49: 230, 1965.
- Cadman, in Virus Diseases of Small Fruits and Grapevines, University of California Press, Berkeley, p. 149, 1970.
- Cadman & Harris, Rep. E. Malling Res. Stn, 1950: 127, 1951.
- Converse, Phytopathology 63: 780, 1973.
- Jones, Rep. Scott. hort. Res. Inst. 1975: in press, 1976.
- Jones & Murant, Rep. Scott. hort. Res. Inst. 1971: 60, 1972a.
- Jones & Murant, Pl. Path. 21: 166, 1972b.
- Lister & Converse, CMI/AAB Descriptions of Plant Viruses: 106, 4 pp.,1972.
- Murant, Proc. Am. Phytopath. Soc. 2: 116, 1975.
- Murant, Chambers & Jones, Ann. appl. Biol. 77: 271, 1974.
- Murant & Jones, Acta Hort. 66: in press, 1976.