Details of DPV and References
DPV NO: 166 September 1976
Family: Tombusviridae
Genus: Carmovirus
Species: Narcissus tip necrosis virus | Acronym: NTNV
Narcissus tip necrosis virus
W. P. Mowat Scottish Horticultural Research Institute, Invergowrie, Dundee, Scotland
C. J. Asjes Bulb Research Centre, Lisse, The Netherlands
A. A. Brunt Glasshouse Crops Research Institute, Littlehampton, Sussex, England
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
Reported by
Asjes (1972) and
Mowat & Hutcheson (1974).
An RNA-containing virus with isometric particles c. 30 nm in diameter which sediment as a single component. Occurs in high concentration in narcissus, the only known host, and is transmissible by manual inoculation. No vector is known. Found in several narcissus cultivars in the Netherlands and UK.
Main Diseases
Found in narcissus cultivars of Royal Horticultural Society's Divisions 1, 2, 3, 4, 8 and 9 (Anon., 1969) and is widespread in at least some stocks of some cultivars. Symptoms appear after flowering. In some cultivars, e.g. Barrett Browning, Goblet and Redmarley, large necrotic lesions appear first near the leaf tips (tip necrosis) (Fig. 1) and then spread basipetally (Fig. 2); in cv. Sempre Avanti the virus induces chlorotic and necrotic streaks mainly in the mid and upper parts of the abaxial leaf surfaces (Fig. 3, Fig. 4). Both kinds of symptom are followed by premature senescence of leaves. Some cultivars, e.g. Fortune and Dutch Master, are symptomlessly infected.
Geographical Distribution
The Netherlands (Asjes, 1972), UK (Mowat & Hutcheson, 1974; Brunt, 1975) and probably wherever commercial cultivars of narcissus are grown.
Host Range and Symptomatology
Narcissus is the only known host. The virus failed to infect plants of 46 other plant species from fourteen families but is readily transmissible to narcissus by inoculation of partially purified preparations. Cultivars differ in the leaf symptoms associated with infection (see Main Diseases). No flower symptoms have been associated with infection.
Strains
Virus isolates that differ slightly in serological tests have been found in naturally infected narcissus but no major strain differences are described. The lack of suitable host species has prevented study of cloned isolates of the virus.
Transmission by Vectors
None known.
Transmission through Seed
No information.
Serology
Antiserum with moderately high titre (1/512) is readily obtained. Double diffusion gel tests give a single band of precipitate and the method is suitable for detecting the virus in crude narcissus sap.
Relationships
Particle size, physical properties and M. Wt of the RNA and protein molecules are similar to those of tombusviruses but no serological relationship was found to five serotypes of tomato bushy stunt virus or to any of 40 other distinct isometric plant viruses, including carnation mottle, carnation ringspot, elderberry latent, pelargonium flower break, saguaro cactus and turnip crinkle viruses.
Stability in Sap
Virus in intact leaves or extracted sap remains infective for at least 7 months at -20°C.
Purification
Virus preparations have been made only from naturally infected narcissus plants, which frequently also contain other viruses. The method used depends on which other viruses are present.
For sources free from other viruses. Extract leaves with neutral 0.017 M phosphate buffer, clarify with an equal volume of diethyl ether and follow with three or more cycles of differential centrifugation, resuspending the virus in the same buffer. This method gives up to 5 mg of virus from 100 g leaf.
For sources also containing filamentous viruses at low concentrations. Extract leaves with neutral 0.067 M phosphate buffer containing 0.1% thioglycollate and freeze for several days at -20°C. Thaw and clarify by adding an equal volume of a 1:1 mixture of n-butanol and chloroform followed by two cycles of differential centrifugation. Freeze the virus suspension at -20°C for 1 h, thaw and centrifuge at low speed.
For sources also infected with narcissus mosaic virus and/or tobacco rattle virus. To sap extracted as in 2) add sodium sulphite-washed n-butanol to 8% (v/v), stir 0.5-1 h, centrifuge at 10,000 g for 20 min and subject the supernatant fluid to one cycle of differential centrifugation, resuspending the virus in 0.03 M phosphate buffer at pH 7.6. Remove tobacco rattle virus by rate zonal centrifugation in 10-40% (w/v) sucrose density gradients and narcissus mosaic virus by exclusion chromatography in columns of 2% agarose beads. Narcissus mosaic virus elutes in the void volume with 0.067 M phosphate buffer at pH 7.6 and narcissus tip necrosis virus is recovered from later fractions by centrifugation.
Properties of Particles
Sediments as a single component with a sedimentation coefficient (s20,w) of 123 S in 0.017 M phosphate buffer at pH 7. Sedimentation coefficients are similar at temperatures from 10-30°C and over the pH range 5-7.
Behaviour in immunoelectrophoresis: In 0.8% Ionagar No. 2 containing 0.017 M phosphate buffer at pH 7 (ionic strength=0.037) or barbitone-acetate buffer at pH 8.6 (ionic strength=0.1) the virus moves as a single component towards the anode.
A260/A280: 1.5-1.6.
Buoyant density: 1.36 g/cm3 in CsCl.
Particle Structure
Particles are isometric (Fig. 7) and about 30 nm in diameter. Particles are disrupted and/or penetrated in 2% potassium phosphotungstate at pH 7, in saturated uranyl formate or in saturated uranyl acetate, but not in 2% potassium phosphotungstate at pH 6 or uranyl formate/NaOH (I. M. Roberts in Barnett & Murant, 1970), or when double stained with ammonium molybdate and uranyl acetate (Home & Ronchetti, 1974) (Fig. 7).
Particle Composition
RNA: M. Wt c. 1.6 x 106, estimated by polyacrylamide gel electrophoresis; approx. 18% of particle weight, estimated from the A260/A280 ratio.
Protein. Two major polypeptide bands of 42,000 and 39,000 daltons M. Wt are obtained by polyacrylamide gel electrophoresis: they are not in equimolar amounts and the smaller possibly represents a degradation product of the larger. The apparent M. Wt of the polypeptides are unaffected by gel concentration.
Relations with Cells and Tissues
In narcissus leaves, virus-like particles occur scattered in the cytoplasm, in cytoplasmic vesicles (Fig. 5) and embedded in a matrix of unknown composition within the vacuoles (Fig. 6, Fig. 8) of cells of the epidermis, mesophyll, vascular parenchyma and phloem (W. P. Mowat & A. M. Hutcheson, unpublished data).
Notes
Eight other viruses with isometric particles c. 30 nm in diameter are known to occur in narcissus: arabis mosaic, broad bean wilt, cucumber mosaic, strawberry latent ringspot, raspberry ringspot, tobacco ringspot, tomato black ring and tomato ringspot. All, unlike narcissus tip necrosis virus, have extensive host ranges and can be detected by inoculation to Chenopodium quinoa. The tip necrosis symptom does not occur in all cultivars and although consistently associated with the virus in some, it may be suppressed by hot dry weather; its use in diagnosis is therefore limited. The virus, however, seems reliably detected by serological double-diffusion gel tests using crude narcissus sap.
The cvs. Barrett Browning and Sempre Avanti are likely to be satisfactory sources of infected plants.
Acknowledgements
Photographs: Figs 1 & 2, Bulb Research Centre; Figs 3-8, Scottish Horticultural Research Institute.
Figures

Tip necrosis in naturally infected narcissus cv. Goblet; early symptom.
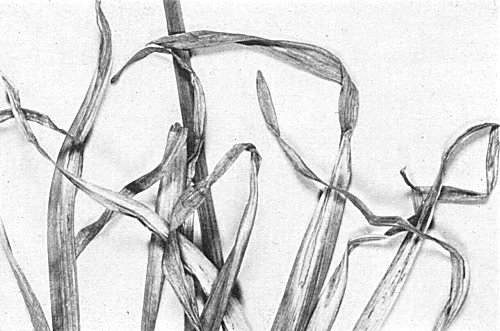
Tip necrosis in naturally infected narcissus cv. Goblet; progressive necrosis.

Chlorotic and necrotic streaks in naturally infected narcissus cv. Sempre Avanti.

Chlorotic and necrotic streaks in naturally infected narcissus cv. Sempre Avanti.

Thin section of infected narcissus leaf cv. Sempre Avanti showing virus particles in the cytoplasm and cytoplasmic vesicles.
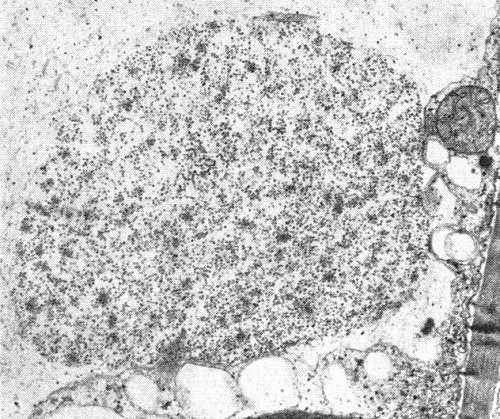
Thin section of infected narcissus leaf cv. Sempre Avanti showing virus particles embedded in a matrix of unknown composition in the vacuole, forming an aggregate.

Particles double stained with ammonium molybdate and uranyl acetate. Bar represents 100 nm.
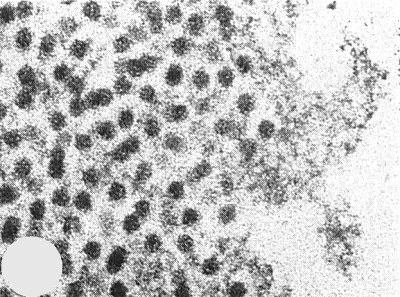
Thin section of infected narcissus leaf cv. Sempre Avanti showing virus particles at the edge of an aggregate: note absence of membrane.
References list for DPV: Narcissus tip necrosis virus (166)
- Anon., Classified List and International Register of Daffodil Names, London: Royal Horticultural Society, 374 pp., 1969.
- Asjes, Jversl. Lab. Bloembollenonderzoek, Lisse, 1970-71: 45, 1972.
- Barnett & Murant, Ann. appl. Biol. 65: 435, 1970.
- Brunt, Rep. Glasshouse Crops Res. Inst., 1974: 116, 1975.
- Home & Ronchetti, J. Ultrastruct. Res. 47: 361, 1974.
- Mowat & Hutcheson, Rep. Scott. hort. Res. Inst. 1973: 61, 1974.