Details of DPV and References
DPV NO: 170 September 1976
Family: Potyviridae
Genus: Macluravirus
Species: Narcissus latent virus | Acronym: NLV
Narcissus latent virus
A. A. Brunt Glasshouse Crops Research Institute, Littlehampton, Sussex, England
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
Detected by Brunt & Atkey (1967)
and partially characterized by
Brunt (1976).
Synonyms
- Irisbontvirus (Asjes & Derks, 1975)
-
A virus with filamentous particles c. 13 x 650 nm; found in W. Europe in narcissus, nerine and bulbous iris, but often present with other viruses. Transmitted in the non-persistent manner by aphids and by mechanical inoculation of sap.
Main Diseases
Infects narcissus, nerine and bulbous iris but the virus is often present with other viruses. In narcissus, the virus alone induces light and dark green mottling near leaf tips of intolerant cultivars (Fig. 1, Fig. 2); this may initially resemble the chlorosis induced by narcissus yellow stripe virus but, unlike the latter symptom, it occurs only on the distal 6-10 cm of leaves. Cultivars with greater tolerance are either symptomless or produce inconspicuous tip chlorosis which is barely discernible through the waxy ‘bloom’ on leaves. The leaf mottling is often more extensive but less conspicuous in infected narcissus plants forced in glasshouses for early flower production. In nerine the virus induces inconspicuous leaf mottling in some cultivars, but is virtually symptomless in others. In bulbous irises it has been found only in complex with other viruses, but in iris seedlings infected by sap-inoculation it induces very faint leaf chlorosis.
Geographical Distribution
Reported in one or more hosts in Britain (Brunt & Atkey, 1967; Brunt, Hollings & Stone, 1970; Brunt, 1976), The Netherlands (Asjes, 1969; Asjes & Derks, 1975) and Germany (Koenig et al., 1973; R. Koenig, personal communication), but probably present wherever nerine, narcissus and bulbous irises are grown.
Host Range and Symptomatology
In addition to the two natural hosts from the Amaryllidaceae and one from the Iridaceae, narcissus latent virus infects 12 of 26 species from the Amaranthaceae, Solanaceae and Papilionaceae but none of 23 species from 11 other families.
-
Diagnostic species
- Tetragonia expansa.
Circular chlorotic lesions in inoculated leaves after 10-14 days, slowly enlarging to 4-8 mm in diameter and often with a green margin (Fig. 3). No systemic infection. - Nicotiana clevelandii, N. megalosiphon and N. benthamiana. Some
isolates induce faint chlorotic lesions in inoculated leaves after 10-15 days but no
systemic infection; others induce both local chlorotic lesions and faint systemic leaf
chlorosis (Fig. 5).
- Phaseolus vulgaris cvs. The Prince and Canadian Wonder. Erratic production of faint chlorotic circular lesions in inoculated primary leaves after 14 days. No systemic infection.
- Narcissus. In intolerant cultivars (e.g. Boswin, Dutch Master) light and dark green leaf mottling mainly near leaf tips (Fig. 1, Fig. 2).
- Phaseolus vulgaris cvs. The Prince and Canadian Wonder. Erratic production of faint chlorotic circular lesions in inoculated primary leaves after 14 days. No systemic infection.
-
Propagation species
- Nicotiana clevelandii
and N. megalosiphon are good sources of inocula and virus for purification, especially of those isolates inducing systemic infection.Assay species
- Although Tetragonia expansa and Phaseolus vulgaris produce local lesions, neither is a reliable assay host. Infectivity is usually best assayed in Nicotiana clevelandii or N. megalosiphon by recording the proportion of inoculated plants becoming infected.
Strains
Although in Nicotiana clevelandii, N. benthamiana and N. megalosiphon some isolates induce local infection only and others infect systemically, all isolates tested are serologically indistinguishable. Such isolates are probably best considered as minor variants rather than as different strains.
Transmission by Vectors
Transmitted efficiently from narcissus and Nicotiana clevelandii in the non-persistent manner by Acyrthosiphon pisum, Aphis gossypii and Myzus persicae.
Transmission through Seed
Not seed-borne in narcissus or Nicotiana clevelandii.
Serology
Antisera with homologous titres of 1/4000 to 1/16,000 are readily prepared in rabbits by one or two intramuscular injections, each of c. 0.5 mg of virus in Freund's complete adjuvant. Antisera react with homologous antigen in tube precipitin tests to produce typical flagellar precipitates.
Relationships
Isolates from narcissus, nerine and bulbous iris are serologically indistinguishable. Antiserum to ‘irisbontvirus’ (Asjes & Derks, 1975) reacted to its homologous titre of 1/3200 with narcissus latent virus. Particle morphology and other properties place narcissus latent virus in the carlavirus group but no relationship was detected to any of fourteen carlaviruses and it is thus probably a distinct member of the group.
Stability in Sap
Sap from Nicotiana clevelandii is usually infective after dilution to 10-3 but not 10-4, after 10 min at 65°C but not 70°C, or after 16 and 4 days at 2°C and 20°C respectively.
Purification
Up to 25 mg virus/kg Nicotiana clevelandii leaf tissue is obtainable by the following procedure (A. A. Brunt & R. J. Barton, unpublished information): Homogenise infected leaves (1 g/2 ml) at 2°C in an extractant at pH 7.1 containing 0.25 M phosphate, 0.2 M ascorbate and 0.1% v/v thioglycollate; add ether and carbon tetrachloride (each 15% v/v), stir the mixture and then subject to one or two cycles of differential centrifugation (15 min at 10,000 g; 80 min at 65,000 g). Resuspend final pellets in 0.033 M phosphate at pH 7.6 (1 ml/50-60 g leaf tissue). Separate the virus from the smaller contaminants (mainly cytoplasmic ribosomes and fraction I protein) by chromatography on 700Å (120-200 mesh) controlled-pore glass beads (Sigma Ltd.); when eluted with 0.05 M phosphate buffer pH 7.6, virus is discharged in or soon after the void volume (Fig. 6) from which it is then readily reconcentrated by ultracentrifugation.
Properties of Particles
Pure virus preparations contain a single sedimenting component with a sedimentation coefficient (s°20, w) of 158±4 S and a buoyant density of 1.33 g/cm3.
A260/A280: 1.09; A260/A240: 1.10 (both values corrected for light-scattering).
Particle Structure
The virus has straight or slightly flexuous filamentous particles c. 13 x 650 nm (Fig. 4).
Particle Composition
Nucleic acid: RNA, probably single-stranded, c. 5 % of particle weight (estimated spectrophotometrically).
Protein: c. 95% of particle weight. Subunit M. Wt (estimated by polyacrylamide gel electrophoresis) c. 32,000 (R. J. Barton & A. A. Brunt, unpublished information).
Relations with Cells and Tissues
No reports.
Notes
The common commercial exchange of flower bulbs throughout Western Europe, together with the detection of the same but no other carlavirus in the three main hosts in Britain, in bulbous iris in Germany (R. Koenig, personal communication) and in narcissus in the Netherlands (Asjes, 1969), all suggested that narcissus latent virus is also the carlavirus infecting nerine and bulbous iris in the Netherlands (Hakkaart, 1972; Asjes & Derks, 1975) and nerine in Germany (Koenig et al., 1973). Although ‘irisbontvirus’ (Asjes & Derks, 1975) and narcissus latent viruses are now known to be synonyms, the serological affinities of nerine latent virus (Hakkaart, 1972) have yet to be determined.
Although the virus is permanently or temporarily symptomless in some narcissus cultivars, in others it induces definite leaf-tip mottling. Nevertheless, the name clearly indicates its affinities with carnation latent and allied viruses (Harrison et al., 1971). The virus alone is probably less damaging than most others infecting the three hosts but, like some other carlaviruses, it probably contributes substantially to the gradual decline of plants when present with other viruses.
The common occurrence of narcissus latent virus and other viruses in the three primary hosts has previously caused difficulties in the separation and identification of the individual viruses, especially when they induced similar symptoms and infected some common indicator plants. Thus local lesions in Tetragonia expansa and systemic chlorosis in Nicotiana clevelandii, which were previously attributed to narcissus yellow stripe virus (Brunt, 1971) or iris mild mosaic virus (Brunt, 1973) respectively, are now known to be induced by narcissus latent virus.
Virus-free nerine plants are readily obtained from infected bulbs by meristem-tip culture; of 250 tips taken from infected plants, 130 produced small plants and of 56 surviving after transplanting into soil, 49 were virus-free (Hakkaart, Maat & Quak, 1975).
Figures

Inconspicuous leaf chlorosis in narcissus cv. Boswin.

Leaf tip chlorosis in narcissus cv. Dutch Master.

Chlorotic local lesions in Tetragonia expansa leaf.

Virus particles in infective Nicotiana clevelandii sap mounted in phosphotungstate. Bar represents 200 nm.

Inconspicuous systemic leaf chlorosis in N. clevelandii.
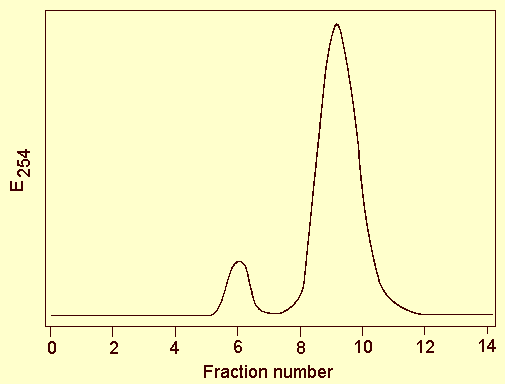
Molecular permeation chromatography elution profile on 700 Å pore size controlled-pore glass beads of a partially purified virus preparation: Fractions 6 and 7 = virus; fractions 9, 10 and 11 = smaller debris.
References list for DPV: Narcissus latent virus (170)
- Asjes, Jversl. Lab. Bloembollen Onderz. Lisse 1968-1969: 40, 1969.
- Asjes & Derks, Jversl. Lab. Bloembollen Onderz. Lisse 1974: 41, 1975.
- Brunt, CMI/AAB Descriptions of Plant Viruses 76: 4 pp., 1971.
- Brunt, CMI/AAB Descriptions of Plant Viruses 116: 4 pp., 1973.
- Brunt, Rep. Glasshouse Crops Res. Inst. 1975: 122, 1976.
- Brunt & Atkey, Rep. Glasshouse Crops Res. Inst. 1966: 155, 1967.
- Brunt, Hollings & Stone, Rep. Glasshouse Crops Res. Inst. 1969: 138, 1970.
- Hakkaart, Jversl. Inst. plziektenk. Onderz. 1971: 105, 1972.
- Hakkaart, Maat & Quak, Acta Hort. 47: 51, 1975.
- Harrison, Finch, Gibbs, Hollings, Shepherd, Valenta & Wetter,Virology 45: 356, 1971.
- Koenig, Lesemann, Brunt & Kühne, Intervirology 1: 346, 1973.