Details of DPV and References
DPV NO: 171 September 1977
Family: Tymoviridae
Genus: Tymovirus
Species: Clitoria yellow vein virus | Acronym: CYVV
Clitoria yellow vein virus
K. R. Bock Ministry of Overseas Development, Crop Virology Project, EAAFRO P.O. Box 30148, Nairobi, Kenya
E. J. Guthrie Ministry of Overseas Development, Crop Virology Project, EAAFRO P.O. Box 30148, Nairobi, Kenya
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
Described by
Bock, Guthrie & Meredith (1977).
An RNA-containing virus with isometric particles c. 28 nm in diameter which sediment as two components. It is readily transmitted by inoculation of sap and infects many species in the Papilionaceae, commonly causing a distinctive vein yellowing. Vector not known. Occurs in Kenya.
Main Diseases
Causes a vein-yellowing disease in Clitoria ternatea (Fig. 1) and a dark green mottle in Abrus precatorius (Bock et al., 1977).
Geographical Distribution
Recorded only from south-eastern (coastal) districts of Kenya, but probably occurs more widely.
Host Range and Symptomatology
Readily transmitted by inoculation of sap to very many species in the Papilionaceae and also to species in the Caesalpiniaceae, Malvaceae and Solanaceae. Commonly causes a distinctive yellowing of secondary and smaller leaf veins. Susceptible genera occur in a wide range of tribes of the Papilionaceae including Abrus (Abreae); Tephrosia (Tephrosieae); Cyamopsis, Indigofera (Indigoferae); Arachis, Stylosanthes (Stylosantheae); Desmodium (Desmodieae); Cajanus, Calopogonium, Canavalia, Centrosema, Cicer, Clitoria, Glycine, Kennedya, Macroptilium, Mucuna, Phaseolus, Vigna, Voandzeia (Phaseoleae); Crotalaria (Genisteae); Medicago, Trifolium, Trigonella (Trifoleae); Hymenocarpus, Securigera (Loteae); Coronilla (Coronilleae); Astragalus (Galegeae); Pisum, Vicia (Vicieae); Hedysarum (Hedysareae) (Bock et al., 1977, and K. R. Bock & E. J. Guthrie, unpublished data).
-
Diagnostic species
- Clitoria ternatea
(butterfly bean, Kordofan pea). Persistent systemic yellow vein chlorosis (Fig. 1). - Phaseolus vulgaris (French bean) cv. Long Tom. Chlorotic local lesions in 6 to
7 days; systemic yellow vein net confined to veinlets in 10 to 11 days
(Fig. 2).
- Nicotiana clevelandii. Persistent systemic whitish vein net in 6 to 7 days (Fig. 3).
- Glycine max (soybean). Systemic yellow vein chlorosis (Fig. 4).
- Bauhinia purpurea. Systemic yellow vein chlorosis (Fig. 5).
- Nicotiana clevelandii. Persistent systemic whitish vein net in 6 to 7 days (Fig. 3).
-
Propagation species
- Bauhinia purpurea
is a satisfactory host for maintaining isolates. Nicotiana clevelandii is a good source for virus purification.Assay species
- Phaseolus vulgaris
(cvs. Long Tom or Premier) is useful for virus assay but no entirely satisfactory local-lesion host is known.
Strains
No evidence of strains. Isolates from Clitoria ternatea and Abrus precatorius are indistinguishable serologically or by host plant reaction.
Transmission by Vectors
No information.
Transmission through Seed
Not seed-borne in Glycine max, Phaseolus vulgaris, Vigna radiata or V. unguiculata.
Transmission by Dodder
No information.
Serology
Strongly immunogenic in rabbits. Antisera with titres of 1/512-1/1024 in gel diffusion tests have been prepared.
Relationships
The virus belongs to the tymovirus group. In agar gel double-diffusion tests the virus reacts with antisera to only four members of the group: it is closely related serologically to cacao yellow mosaic and kennedya yellow mosaic, and more distantly related to desmodium yellow mottle and okra mosaic viruses (Bock et al., 1977). The interrelationships among these five viruses can be expressed diagrammatically as a ‘loop’ structure (Koenig, 1976). Clitoria yellow vein virus has many properties typical of the group including particle morphology, sedimentation behaviour, molecular weight of the coat polypeptide, high nucleic acid content, high thermal inactivation point and high concentration in host plants.
Stability in Sap
In N. clevelandii sap, the dilution end-point is about 10-8, the thermal inactivation point about 72°C, and infectivity is retained for about 3 weeks at 18-20°C.
Purification
(Bock et al., 1977). Homogenise infected N. clevelandii leaves in 0.1 M sodium borate buffer containing 0.001 M ethylene diamine tetraacetic acid and 1% 2-mercaptoethanol, pH 8.5 (2 ml/g tissue). Clarify with an equal volume of a 1:1 mixture of n-butanol and chloroform; concentrate the virus by two or three cycles of differential centrifugation, resuspending pellets in 0.01 M phosphate buffer (pH 7.3). Further purification and separation of top and bottom components may be achieved by sucrose density gradient centrifugation.
Properties of Particles
As with other tymoviruses, there are two classes of particles of similar diameter which sediment as two components in the ultracentrifuge (Fig. 6) and in sucrose density gradient centrifugation. The top component (T) consists of protein shells without nucleic acid, the bottom component (B) contains infective nucleoprotein particles.
Sedimentation coefficient, (s20, w) (svedbergs): 50(T) and 109(B) at infinite dilution.
A260/A280: 1.69(B).
Amax/Amin: 1.46(B).
Particle Structure
Particles are isometric, c. 28 nm in diameter (Fig. 7); T component particles are penetrated by 2% sodium phosphotungstate, whereas B component particles are not.
Particle Composition
Nucleic acid: probably RNA, single-stranded, about 37% of particle weight (estimated according to Reichmann, 1965).
Protein: M. Wt c. 20,000 by SDS-polyacrylamide gel electrophoresis.
Relations with Cells and Tissues
No information.
Notes
Clitoria yellow vein virus may be distinguished from two other African tymoviruses, cacao yellow mosaic and okra mosaic, by its inability to infect species of Cucurbitaceae and Chenopodiaceae. With the possible exception of cacao yellow mosaic virus, it is readily distinguishable from all other tymoviruses by double-diffusion serological tests in agar gel.
Figures

Systemic symptoms in Clitoria ternatea.
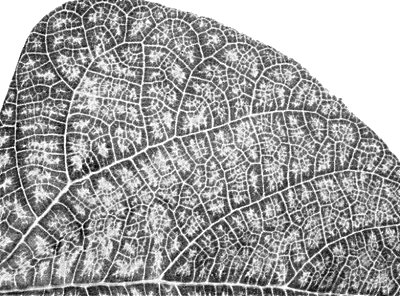
Systemic symptoms in Phaseolus vulgaris cv. Long Tom.

Systemic symptoms in Nicotiana clevelandii.

Systemic symptoms in Glycine max.

Systemic symptoms in Bauhinia purpurea.
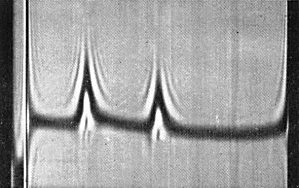
Schlieren pattern obtained by analytical centrifugation showing (left) 50 S and (right) 109 S components. (Photo courtesy R. D. Woods.)

Purified virus preparation in 2% sodium phosphotungstate. Bar represents 100 nm. (Photo courtesy R. D. Woods.)
References list for DPV: Clitoria yellow vein virus (171)
- Bock, Guthrie & Meredith, Ann. appl. Biol. 85: 97, 1977.
- Koenig, Virology 72: 1, 1976.
- Reichmann, Virology 25: 166, 1965.