Details of DPV and References
DPV NO: 175 September 1977
Family: Reoviridae
Genus: Fijivirus
Species: Pangola stunt virus | Acronym: PaSV
Pangola stunt virus
R. G. Milne Laboratorio di Fitovirologia applicata del CNR, 10135 Torino, Italy
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
First described by
Dirven & van Hoof (1960) and
Schank & Edwardson (1968).
A virus with polyhedral particles 65-70 nm in diameter, having a double-shelled capsid and containing double-stranded RNA. Host range confined to Gramineae and its planthopper (Delphacidae) vector. Not mechanically transmissible to plants. Causes a severe disease of pangola grass.
Main Diseases
Causes severe stunting of the tropical herbage plant pangola grass (Digitaria decumbens) and poor regeneration after cropping. Other symptoms include yellowing, kinking or twisting of young leaves and inflorescences (Fig. 1, Fig. 2), small vein swellings, excess tillering, and purpling of leaf margins. The darker green colour induced in grasses by several related viruses does not occur. The incidence of pangola stunt is highest in the dry season and in dry years, probably because these conditions favour the vector (Dirven & van Hoof, 1960; Hunkar et al., 1974).
Geographical Distribution
Guyana, Brazil, Peru, Fiji and Taiwan.
Host Range and Symptomatology
Only plants within the genus Digitaria have been investigated. It is likely that other genera within the Gramineae will prove susceptible. There is great variability in field resistance not only between different species of Digitaria but also between different lines within species (Hunkar et al., 1974). The planthopper Sogatella furcifera (Horvath) is the only reported insect host. No disease is reported in the insect. The virus is not transmissible to plants by inoculation of sap.
-
Diagnostic species
- Highly susceptible species and varieties of Digitaria, e.g. pangola grass (D. decumbens, line PI 111110), D. longiflora 299642, D. pentzii 299769, D. setivalvola 299795, D. valida 299875 and others (see Hunkar et al., 1974). The diagnostic symptoms are those described under Main Disease. In the glasshouse, small enations detectable with a lens occur. In the field, some cells in the sheaths of the vascular bundles acquire extremely thick walls which may be diagnostic (Schank et al., 1972).
-
Propagation species
- Probably any highly susceptible clone of Digitaria. Pangola grass itself has been used. Infected plants can be increased vegetatively.
-
Assay species
- None tested, but the lines listed above may be used for assaying transmission by vectors. No local lesion host.
Strains
None reported.
Transmission by Vectors
The only known vector is the planthopper Sogatella furcifera (Delphacidae). Incubation in the vector is said to take 2-3 months (Dirven & van Hoof, 1960). Thin sectioning and electron microscopy show that the virus multiplies in nearly all vector tissues (Kitajima & Costa, 1970). Transmission through the egg has not been tested nor have other details of transmission been reported.
Transmission through Seed
No reports (pangola grass itself is a sterile triploid).
Transmission by Dodder
No reports.
Serology
The B-spiked inner shell of the capsid (see Particle structure) is a good immunogen but the outer shell and A spikes are very poor immunogens and there are no data about them. Antiserum prepared by injecting rabbits in the foot pads with partially purified B-spiked inner shells reacted at dilutions up to 1/256 in double diffusion tests with the same preparation but not with healthy Digitaria proteins. Gel diffusion and immune electron microscopy have shown that the B spikes and the inner shell but not the outer shell react with this antiserum. The double-stranded RNA of the virus is weakly immunogenic (Milne & Luisoni, 1977).
Relationships
Pangola stunt virus is in the reovirus group, sharing many properties with other mammalian, avian, insect and plant reo-like viruses (Reoviridae) (Fenner, 1976). The virus is distinct from wound tumor and rice dwarf viruses in capsid structure (unlike these, it has spikes on the outer shell and easily loses parts of the outer shell to give a spiked inner shell), in genome pattern (it has 9 or 10 genome segments, not 12) and in vector type (Delphacid, not Cicadellid). In these respects it is similar to maize rough dwarf and rice black streaked dwarf viruses. However, it differs from these in having a different vector within the Delphacidae, a much longer incubation period in the vector, in inducing a yellow rather than a dark green colour in infected plants and in failing to produce conspicuous enations on infected leaves. Antigenically, the spiked inner shell is similar but not identical to that of maize rough dwarf virus.
In capsid and vector it is also similar to several viruses whose genomes are undescribed: cereal tillering disease virus, oat sterile dwarf virus, arrhenatherum blue dwarf virus and lolium enation virus (Milne & Lovisolo, 1977; G. Boccardo, unpublished data).
Serologically, the inner shell and B spikes are fairly closely related to those of maize rough dwarf virus (Fig. 5, Fig. 6), rice black streaked dwarf virus and cereal tillering disease virus, but apparently unrelated to those of sugarcane Fiji disease virus, oat sterile dwarf virus, arrhenatherum blue dwarf virus and lolium enation virus (Milne & Luisoni, 1977).
Stability in Sap
Not investigated.
Purification
Only partial purification of the B-spiked inner shells has been achieved. Roots and stem bases of infected pangola grass are crushed in 0.05 M phosphate buffer, pH 7, in the cold. The extract is shaken with Freon 113 (1,1,2-trifluoro-1,2,2-trichloroethane) and the upper phase ultracentrifuged into a sucrose density gradient. The zone containing spiked inner shells (sometimes visible as a light-scattering band) is withdrawn, diluted and pelleted, to be resuspended in buffer. Alternatively, leaves are homogenised in buffer as above and the extract is shaken with chloroform, followed by two or three cycles of differential centrifugation.
Properties of Particles
Virus particles are unstable in 2% neutral phosphotungstate, 1 M NaCl, 1 M CsCl, chloroform or n-butanol, the outer shell being stripped to leave spiked or smooth inner shells. Other properties not investigated.
Particle Structure
In uranyl acetate negative stain, with or without previous glutaraldehyde fixation, the virus particle appears spherical or slightly angular, with a double shell and icosahedral symmetry. It is about 65 nm in diameter, with an apparently solid core (Fig. 10). It possesses 12 projections (A spikes) about 10 nm long and 10 nm wide. Morphological subunits are discernible in the outer shell but their number and arrangement are uncertain. The A spikes and parts of the outer shell are easily lost to give a spherical inner shell about 50 nm in diameter possessing 12 hollow B spikes 8 nm long and 12 nm wide (Fig. 11). The B spikes can be lost to give a smooth inner shell (Fig. 12). In thin sections, the virus particle appears round or angular, 65-68 nm in diameter, with a dense core about 50 nm in diameter (Fig. 4). Spiked inner shells are also seen. In negative stain and in thin sections, the particles are indistinguishable from those of maize rough dwarf and a number of similar viruses (Giannotti & Milne, 1977).
Particle Composition
-
Nucleic acid: The inner shell contains double-stranded RNA. The
pattern of the genome bears a close similarity to that of
maize rough dwarf virus.
However, the only isolate investigated, cultured without transmission
by the vector for several years, has 9 linear segments rather than the 10
expected: the segment corresponding to No. 8 of maize rough dwarf virus is
absent or very faint after electrophoresis in polyacrylamide gels
(Fig. 9).
In
phosphate-Tris-EDTA buffer in 5% gels, and taking the maize rough dwarf virus
genome as standard
(Reddy et al., 1975),
the genome segments of pangola
stunt virus have the following estimated M. Wt (the figure in parenthesis refers
to the faint zone corresponding to segment No. 8 of maize rough dwarf virus):
2.92, 2.54, 2.50, 2.50, 2.12, 1.75, 1.48, (1.25), 1.18, 1.12 (all x 106
daltons) (G. Boccardo, unpublished data).
Protein: No information.
Relations with Cells and Tissues
Numerous virus particles occur in the cytoplasm of phloem parenchyma and young sieve tubes, individually or in crystalline array (Schank & Edwardson, 1968; Kitajima & Costa, 1970; Schank et al., 1972; Giannotti & Milne, 1977). ‘Viroplasms’ up to 10 µm in diameter occur in infected cells (Fig. 7). These consist of filamentous matrices in which are embedded empty and full inner shells (Fig. 3). Virus particles may lie in single rows inside straight tubes 100-120 nm in diameter (Fig. 8, Fig. 4). The tubes may take an open or scroll-like form. Mitochondria, chloroplasts and nuclei are not primarily affected by the virus. In field plants, some of the bundle sheath cells become extremely thick-walled.
The virus multiplies in most insect tissues. The cytopathology, including the appearance of virus particles, viroplasms and tubes, is similar to that in plants (Kitajima & Costa, 1970).
Notes
Pangola stunt virus may be identified using immune electron microscopy on crude sap from roots or stem bases of suspect plants, or using gel double diffusion methods on concentrated sap extracts, with antisera against pangola stunt virus, maize rough dwarf virus or rice black streaked dwarf virus (Milne & Luisoni, 1977). Serological detection of the double-stranded RNA should also be possible (Ikegami & Francki, 1973).
The best means of controlling pangola stunt appears to be to find or develop resistant or tolerant lines of Digitaria. Boyd et al. (1973) report one such promising line from South Africa, D. decumbens ‘Transvaal digitgrass’, PI 299601.
Figures

Pangola grass inflorescences, (left) healthy, (middle and right) infected.

Pangola grass leaves, (below) healthy, (above) infected.

Thin section through disintegrating viroplasm in infected pangola grass cell, showing filaments and empty and full inner shells. Bar represents 200 nm. (Courtesy of Dr J. Giannotti.)
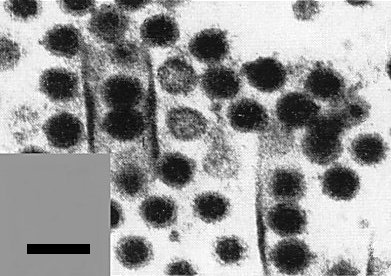
Virus particles and tubular structures in a thin section of an infected pangola grass cell. Bar represents 100 nm. (Courtesy of Dr J. Giannotti.)

Immunodiffusion reactions of pangola stunt (P) and maize rough dwarf (M) B-spiked inner shells with maize rough dwarf virus anti-inner shell (c) serum. (Courtesy of Dr E. Luisoni.)

Immunodiffusion reactions of pangola stunt (P) and maize rough dwarf (M) B-spiked inner shells with maize rough dwarf virus anti-B spike (s) serum. (Courtesy of Dr E. Luisoni.)

Thin section through infected pangola grass phloem parenchyma, showing virus crystals, viroplasm (V), empty inner shells (I) and nucleus (N). Bar represents 1 µm. (Courtesy of Dr J. Giannotti.)

A tube containing virus particles, in phosphotungstate after fixation in glutaraldehyde. Bar represents 100 nm.
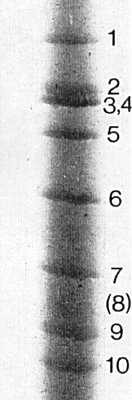
The double-stranded RNA genome segments after electrophoresis in 5% polyacrylamide phosphate-Tris-EDTA buffer. Bands 3 and 4 are superimposed. (Courtesy of Dr G. Boccardo.)
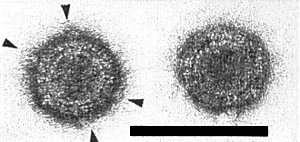
Virus particles in uranyl acetate (UA). A spikes are arrowed. Bar represents 100 nm.
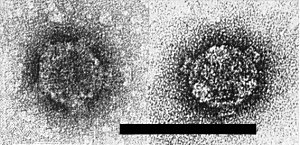
B-spiked inner shells in UA, showing 5-fold and 3-fold symmetries. Bar represents 100 nm.
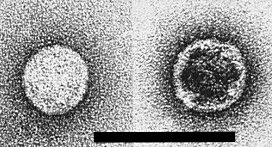
Smooth inner shells in UA. Bar represents 100 nm.
References list for DPV: Pangola stunt virus (175)
- Boyd, Schank, Smith, Hodges, West, Kretschmer, Brolmann & Moore, Circ. Fla Univ. agric. Exp. Stn No. S-222, 16 pp., 1973.
- Dirven & van Hoof, Tijdschr. PlZiekt. 66: 344, 1960.
- Fenner, Intervirology 7: 1, 1976.
- Giannotti & Milne, Virology 80: 347, 1977.
- Hunkar, Hung, Dulder, Schank, Holder & Edwards, Tropical Agriculture 52: 75, 1974.
- Ikegami & Francki, Virology 56: 404, 1973.
- Kitajima & Costa, In 7th International Congress of Electron Microscopy, Vol. 3, p. 323, Ed. P. Favard, 1970.
- Milne & Lovisolo, Adv. Virus Res. 21: 267, 1977.
- Milne & Luisoni, Virology 80: 12, 1977.
- Reddy, Boccardo, Outridge, Teakle & Black, Virology 63: 287, 1975.
- Schank & Edwardson, Crop Sci. 8:118, 1968.
- Schank, Edwardson, Christie & Overman, Euphytica 21: 344, 1972.