Details of DPV and References
DPV NO: 187 August 1978
Family: Betaflexiviridae
Genus: Tepovirus
Species: Potato virus T | Acronym: PVT
Potato virus T
L. F. Salazar Centro Internacional de la Papa, Apartado 5969, Lima, Peru
B. D. Harrison Scottish Horticultural Research Institute, Invergowrie, Dundee, Scotland
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
Described by
Salazar & Harrison (1977,
1978).
An RNA-containing virus with flexuous filamentous particles c. 640 x 12 nm. It has a somewhat limited host range, is seed-borne in some solanaceous species and is readily transmitted by inoculation with sap.
Main Diseases
Reported only from potato (Solanum tuberosum) in which it is usually latent, but occasionally produces a mild leaf mottle.
Geographical Distribution
Recorded only from Peru (Salazar, 1972) and Bolivia (J. Abad & C. Fribourg, unpublished results), but probably occurs more widely in the Andean region of S. America.
Host Range and Symptomatology
Host range is somewhat limited; 46 species in 8 dicotyledonous families are susceptible to infection (Salazar & Harrison, 1978). Most susceptible species are in the Solanaceae, Leguminosae and Chenopodiaceae.
-
Diagnostic species
- Chenopodium amaranticolor.
No symptoms in inoculated leaves. Systemic necrosis develops about 8 days after inoculation (Fig. 1); subsequently plants produce less severely affected leaves that contain virus. - C. quinoa. Inoculated leaves are symptomless or develop chlorotic spots.
Systemic mosaic is produced in plants grown in high intensity light
(Fig. 2)
and tip
necrosis in low intensity light.
- Datura stramonium. Inoculated leaves are symptomless. Systemically infected leaves develop mild mosaic in 8-10 days.
- Nicotiana debneyi. No symptoms in inoculated leaves. Systemically infected leaves show mottling and some necrosis (Fig. 3).
- Phaseolus vulgaris (French bean) cvs Pinto and The Prince. Necrotic local lesions in leaves shaded heavily after inoculation (Fig. 4). Systemic necrosis and recovery of plants.
- Datura stramonium. Inoculated leaves are symptomless. Systemically infected leaves develop mild mosaic in 8-10 days.
-
Propagation species
- Chenopodium quinoa
is suitable as a source of virus for purification.Assay species
- Phaseolus vulgaris
cv. Pinto or The Prince can be used for local lesion assay. Chenopodium amaranticolor is useful for indexing and as a systemic host.
Strains
An isolate differing in virulence has been obtained from Bolivia (J. Abad & C. Fribourg, unpublished results).
Transmission by Vectors
None known; not transmitted by the aphids Myzus persicae or Macrosiphum euphorbiae (Salazar & Harrison, 1978).
Transmission through Seed
Frequently seed transmitted in Datura stramonium (up to 72%), Nicandra physalodes (28%), and Solanum demissum A (39%) and Y (10%). Also transmitted from pollen to seed but not to pollinated plants of Solanum demissum A. Not pollen transmitted in D. stramonium and N. physalodes (Salazar & Harrison, 1978).
Serology
Moderately immunogenic. Antiserum with a precipitin titre of 1/512 was obtained. Virus in clarified or partially purified virus preparations can be detected by tube precipitin tests. Enzyme-linked immunosorbent assay and electron microscopy of virus + antiserum mixtures can also be used for detection and diagnosis.
Relationships
The virus is serologically related to apple stem grooving virus but not to apple chlorotic leaf spot, potato S, potato M, potato Y or potato X viruses. In Chenopodium quinoa, a mixed inoculum containing potato virus T and apple stem grooving virus caused more severe symptoms than did inoculum containing either virus alone, suggesting that the two viruses are not closely enough related to interfere with each other’s multiplication (Salazar & Harrison, 1978). Although they share some properties with closteroviruses, potato T and apple stem grooving viruses should probably not be included in this group.
Stability in Sap
In Chenopodium quinoa sap, the thermal inactivation point (10 min) is about 65°C, dilution end-point about 10-5, and infectivity is retained at 20°C for 2-4 days (Salazar & Harrison, 1977).
Purification
Extracts of systemically infected Chenopodium quinoa leaves can be clarified with 8.5% butanol but larger virus yields are obtained when they are clarified with bentonite as described by Lister (1971) for apple stem grooving virus. The following method (Salazar & Harrison, 1978) is satisfactory. Triturate tissue in 0.06 M phosphate buffer, pH 7.0 (2:1, w/v), filter, and shake filtrate with 3-4% bentonite suspension (c. 50 mg/ml in phosphate buffer; bentonite fraction is that sedimenting between 3000 and 10,000 g). Clarify by low speed centrifugation. Repeat treatment with small additions of bentonite until extract is straw yellow in colour. Precipitate virus with 5-6% polyethylene glycol for 1 h at 4°C. Re-suspend sediment in the buffer, and purify the virus further by differential centrifugation and sucrose gradient centrifugation. The yield is 3-10 mg virus per kg leaf tissue.
Properties of Particles
Purified preparations contain a single sedimenting component observed by analytical or density gradient centrifugation.
Sedimentation coefficient (s20,w) at infinite dilution: 99 S.
A260/A280: 1.18.
Particle Structure
Particles are flexuous filaments c. 637 x 12 nm (Fig. 6). They are helically constructed with a pitch of 3.4 nm and between 9 and 10 subunits per turn of the basic (primary) helix. They are partially degraded in 1-2% sodium phosphotungstate or 1-2% ammonium molybdate. In uranyl formate, the particles are very flexuous and show cross-banding, whereas in uranyl acetate they are straighter and show two types of substructural detail, called ‘criss-cross’ (Fig. 5) and ‘rope’, produced by clustering of the protein subunits (Salazar et al., 1978). Apparently degraded virus particles called ‘ghosts’ are seen in some purified preparations of virus particles. Disc aggregates of virus particle protein, which are 12 nm in diameter, can be produced when virus nucleoprotein particles are disrupted (L. F. Salazar & B. D. Harrison, unpublished results).
Particle Composition
Nucleic acid: Single-stranded RNA of M. Wt c. 2.2 x 106, estimated by electrophoresis at 60°C in polyacrylamide gels containing 8 M urea; about 5% of particle weight (Salazar & Harrison, 1978).
Protein: Subunits have M. Wt of about 27,000, estimated by SDS/polyacrylamide gel electrophoresis (5% to 10.5% gels).
Relations with Cells and Tissues
Not studied.
Notes
Unlike other potato viruses with filamentous particles, potato virus T infects Phaseolus vulgaris systemically. Apple stem grooving virus, although serologically related to potato virus T, apparently does not infect potato (Salazar & Harrison, 1978). None of these viruses has particles that show the criss-cross patterns typical of potato virus T when stained with uranyl acetate.
Acknowledgements
Photographs: after Salazar & Harrison, 1978.
Figures

Systemic necrosis in Chenopodium amaranticolor.

Systemic mild mosaic in C. quinoa.

Systemic mottle in Nicotiana debneyi leaf.
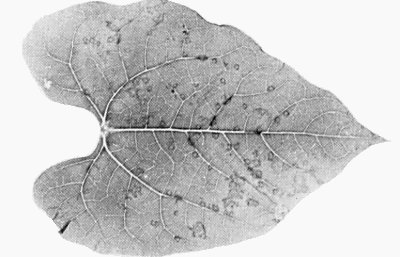
Local lesions in inoculated leaf of Phaseolus vulgaris cv. The Prince.

Virus particle stained with uranyl acetate, showing ‘criss-cross’ pattern. Bar represents 100 nm.

Virus particles stained with sodium phosphotungstate at pH 7. Bar represents 100 nm.
References list for DPV: Potato virus T (187)
- Lister, Annls Phytopath. Numéro hors série: 363, 1971.
- Salazar, Investnes agropec. Peru 3: 3, 1972.
- Salazar & Harrison, Nature, Lond. 265: 337, 1977.
- Salazar & Harrison, Ann. appl. Biol. 89: 223, 1978.
- Salazar, Hutcheson, Tollin & Wilson, J. gen. Virol. 39: 333, 1978.