Details of DPV and References
DPV NO: 191 August 1978
Family: Potyviridae
Genus: Potyvirus
Species: Dasheen mosaic virus | Acronym: DsMV
Dasheen mosaic virus
F. W. Zettler Plant Virus Laboratory, Plant Pathology Department, University of Florida, Gainesville, Florida 32611, USA
M. M. Abo El-Nil Forestry Research Center, Weyerhaeuser Corporation, 505 N. Pearl Street, Centralia, Washington 98531, USA
R. D. Hartman P.O. Box 90, Palmdale, Florida 33944, USA
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
- Described by Zettler et al. (1970) and Abo El-Nil, Zettler & Hiebert (1977).
- A virus with flexuous, filamentous particles c. 750 nm long. It is transmitted by aphids in the non-persistent manner, is sap-transmissible, and infects members of the family Araceae.
Main Diseases
Causes mosaic and/or distortion diseases of Aglaonema spp., Caladium hortulanum (caladium), Colocasia spp. (dasheen, taro), Dieffenbachia spp. (dieffenbachia, dumbcane), Xanthosoma spp. (cocoyam, elephant’s ear, tannia, yautia) and Zantedeschia spp. (calla lily).
Geographical Distribution
Appears to be world-wide, especially in tropical and subtropical regions. Reported from the Caribbean (Alconero & Zettler, 1971; Debrot & Ordosgoitti, 1974; Hartman, 1974a), Egypt (Abo El-Nil & Zettler, 1976), Florida, USA (Hartman & Zettler, 1972; Zettler et al., 1970), Europe (Hakkaart & Waterreus, 1976; Van Hoof, 1972; Zettler et al., 1970), India (Hartman, 1974a), Japan (Arai, Doi & Yora, 1970) and Oceania (Alconero & Zettler, 1971; Buddenhagen, Milbrath & Hsieh, 1970; Gollifer et al., 1977; Kenten & Woods, 1973).
Host Range and Symptomatology
Reported to infect species of the following 13 genera in the family Araceae:
Aglaonema,
Alocasia, Amorphophallus, Anthurium, Arisaema, Caladium, Colocasia, Cryptocoryne, Dieffenbachia,
Philodendron, Spathiphyllum, Xanthosoma, Zantedeschia (Hartman, 1974a; Hartman &
Zettler, 1972; Tooyama, 1975a; Zettler et al., 1970).
- Diagnostic species
- Philodendron selloum
. Young seedlings show systemic vein chlorosis c. 2-3 weeks after inoculation (Fig. 1). This symptom is usually followed by severe mosaic and deformation of subsequently formed leaves (Fig. 2). - Caladium hortulanum, Colocasia esculenta or Xanthosoma caracu. Foliar mosaic
symptoms, often localized. Mosaic patterns may appear as conspicuous localized ‘feathering’
along the veins (Fig. 3). Many leaves of infected plants may be symptomless (Tooyama, 1975b;
Zettler et al., 1970).
- Dieffenbachia picta. Leaves show mosaic and distortion symptoms. In some instances, the unfurling leaf may be distorted. Mature leaves may be torn due to uneven expansion rates of laminar tissues. Leaves may be symptomless during certain parts of the growing season (Hakkaart & Waterreus, 1976; Hartman, 1974a).
- Zantedeschia. Leaves show mosaic and distortion symptoms.
- Dieffenbachia picta. Leaves show mosaic and distortion symptoms. In some instances, the unfurling leaf may be distorted. Mature leaves may be torn due to uneven expansion rates of laminar tissues. Leaves may be symptomless during certain parts of the growing season (Hakkaart & Waterreus, 1976; Hartman, 1974a).
- Propagation species
- Philodendron selloum
. Seedling plants are useful as a source for virus purification and for maintaining virus cultures.- Assay species
- Philodendron verrucosum
is reported to be a local lesion host (Tooyama, 1975a). Philodendron selloum is a useful test plant for aphid transmission tests.
Strains
None distinguished, although minor differences in symptoms induced by single isolates from Florida and Fiji in Philodendron selloum have been noted (Abo El-Nil et al., 1977).
Transmission by Vectors
Transmitted in a non-persistent manner by aphids including Myzus persicae and Aphis craccivora, but not apparently by Pentalonia nigronervosa (Morales & Zettler, 1977).
Transmission through Seed
None detected. Seedlings of Caladium hortulanum, Dieffenbachia picta and Xanthosoma caracu derived from infected parental stock were healthy (Hartman et al., 1972; Volin & Zettler, 1976).
Serology
The virus is a good immunogen. It reacts well in immunodiffusion tests conducted in agar gels containing sodium dodecyl sulphate (Fig. 4) or in agar gels after pyrrolidine treatment of the antigen (Abo El-Nil et al., 1977). The proteinaceous cylindrical inclusions are also immunogenic, are detectable in agar gels containing sodium dodecyl sulphate, and are immunochemically distinct from viral coat protein (F. J. Morales, F. W. Zettler & E. Hiebert, unpublished data).
Relationships
This virus has been classified in the potyvirus group on the basis of particle morphology, aphid-transmissibility, ability to induce cylindrical (‘pinwheel’) inclusions in its hosts, and serological relationships to other potyviruses (Abo El-Nil et al., 1977; Zettler et al., 1970). Immunodiffusion tests conducted in agar gels containing sodium dodecyl sulphate indicate that this virus is related to blackeye cowpea mosaic and tobacco etch viruses (Abo El-Nil et al., 1977).
Stability in Sap
In sap from infected Philodendron selloum, the virus was inactivated by heating to 60-65°C for 10 min by dilution beyond 10-2, or by storage for more than 75 h at 26°C (Alconero, 1972).
Purification
The virus has been purified from systemically infected leaves of Philodendron selloum harvested 2-3 weeks after inoculation. Homogenise tissue (100 g) in a chilled mixture of 200 ml 0.1 M sodium citrate (pH 7.2) containing 0.6 g sodium sulphite and 0.01 M disodium ethylene diamine tetraacetate, 45 ml chloroform, and 45 ml carbon tetrachloride. Centrifuge the homogenate at 13,200 g for 10 min. Precipitate the virus from the supernatant fluid by adding polyethylene glycol M. Wt 6000 to 8% (w/v) and purify further using differential centrifugation and equilibrium centrifugation in CsCl (Abo El-Nil et al., 1977).
Virus-induced cylindrical inclusions of this virus can be obtained from the tissue extract after treatment with chloroform and carbon tetrachloride (F. J. Morales, F. W. Zettler & E. Hiebert, unpublished data). Clarify the homogenate by centrifugation at 4000 g for 5 min then centrifuge the supernatant fluid at 13,200 g for 15 min. Resuspend the resulting pellet in 0.02 M phosphate (pH 8.2) containing 0.1% mercaptoethanol and add Triton X-100 to 5% (v/v) and stir for 1 h. To remove the detergent, use two cycles of centrifugation at 20,000 g for 20 min. Resuspend the pellet and further purify the inclusions by sucrose density gradient centrifugation (Hiebert & McDonald, 1973).
Properties of Particles
Absorbance at 260 nm (1 mg/ml, 1 cm light path): 2.38.
A260/A280: 1.09-1.19.
Particle Structure
Particles are flexuous filaments (Fig. 5), about 750 nm long (Zettler et al., 1970).
Particle Composition
Nucleic acid: No information.
Protein: Electrophoresis of virus coat protein in 11% polyacrylamide gels containing sodium dodecyl sulphate revealed up to six components ranging in estimated M. Wt from 25,000 to 45,000. The major components, the two largest resolved, showed anomalous electrophoretic behaviour, giving much higher estimates of M. Wt in 3% and 6% than in 11% gels (Abo El-Nil et al., 1977).
Relations with Cells and Tissues
The virus induces cytoplasmic cylindrical inclusions which can be detected by light microscopy (Fig. 6) and electron microscopy (Fig. 7, Fig. 8) (Abo El-Nil & Zettler, 1976; Zettler et al., 1970) The cylindrical inclusion protein subunit has a M. Wt of c. 69,000 (F. J. Morales, F. W. Zettler & E. Hiebert, unpublished data).
Notes
Filamentous viruses infecting Anthurium andraeanum in Venezuela (Herold, 1967) and Zantedeschia aethiopica in the Soviet Union (Kolbasina & Protsenko, 1973) have been reported, but their relationship to dasheen mosaic virus has not been determined. Cucumber mosaic virus (Lovisolo & Conti, 1969), tomato spotted wilt virus (Tompkins & Severin, 1950) and two bacilliform viruses infecting taro (James, Kenten & Woods, 1973) can readily be distinguished from dasheen mosaic virus by differences in particle morphology, host range and/or mode of transmission.
The widespread incidence of dasheen mosaic virus reported in plantings of Aglaonema, Caladium, Colocasia, Dieffenbachia, Xanthosoma and Zantedeschia (Abo El-Nil & Zettler, 1976; Alconero & Zettler, 1971; Debrot & Ordosgoitti, 1974; Hartman & Zettler, 1972; Zettler et al., 1970) probably results from its transmissibility by aphids and the use of vegetative means for propagating these plants. Tissue culture techniques have been used to obtain virus-free plants of Caladium, Colocasia, and Xanthosoma (Hartman, 1974b). Attempts to eliminate dasheen mosaic virus from Xanthosoma corms by heat treatment were unsuccessful (Alconero, 1972).
Figures

Chlorosis of Philodendron selloum leaf.

Mosaic and distortion of P. selloum leaf.

Mosaic of taro (Colocasia esculenta) leaf.

Immunodiffusion test showing the serological relationship of dasheen mosaic virus to blackeye cowpea mosaic virus. Centre well contains antiserum to dasheen mosaic virus. Peripheral wells contain extracts of: (a) P. selloum infected with dasheen mosaic virus, (b) cowpea (Vigna unguiculata) infected with blackeye cowpea mosaic virus, (c) healthy cowpea, and (d) healthy P. selloum. The diffusion medium contains 0.5% sodium dodecyl sulphate, and non-degraded antigens were tested.
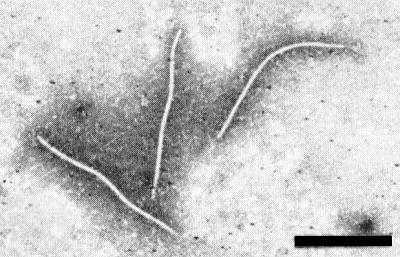
Filamentous virus particles mounted in phosphotungatate. Bar represents 500 nm.

Photomicrograph of cylindrical inclusions (arrows) in epidermal cell of taro. Bar represents 5 µm. (Photomicrograph courtesy of R. G. Christie.)

Electron micrograph of cylindrical inclusions in cell of calla lily (Zantedeschia elliotiana). Bar represents 500 nm.

Cylindrical inclusions in leaf extract of Egyptian taro (Colocasia antiquorum) in ammonium molybdate. Bar represents 250 nm. Inset is an enlarged portion of a cylindrical inclusion (plate) showing striations (periodicity is 5 nm).
References list for DPV: Dasheen mosaic virus (191)
- Abo El-Nil & Zettler, Pl. Dis. Reptr 60: 281, 1976.
- Abo El-Nil, Zettler & Hiebert, Phytopathology 67: 1445, 1977.
- Alconero, Pl. Dis. Reptr 56: 320, 1972.
- Alconero & Zettler, Pl. Dis. Reptr 55: 506, 1971.
- Arai, Doi & Yora, Ann. Phytopath. Soc. Japan 36: 363, 1970.
- Buddenhagen, Milbrath & Hsieh, Proc. int. Symp. trop. Root & Tuber Crops 2: 53, 1970.
- Debrot & Ordosgoitti, Pl. Dis. Reptr 58: 1032, 1974.
- Gollifer, Jackson, Dabek, Plumb & May, PANS 23: 171, 1977.
- Hakkaart & Waterreus, Acta Hort. 59: 175, 1976.
- Hartman, Ph.D. Dissertation, Univ. Fla, 1974a.
- Hartman, Phytopathology 64: 237, 1974b.
- Hartman & Zettler, Phytopathology 62: 804, 1972.
- Hartman, Zettler, Knauss & Hawkins, Proc. Fla St. hort. Soc. 85: 404, 1972.
- Herold, Phytopathology 57: 8, 1967.
- Hiebert & McDonald, Virology 56: 349, 1973.
- James, Kenten & Woods, J. gen. Virol. 21: 145, 1973.
- Kenten & Woods, PANS 19: 38, 1973.
- Kolbasina & Protsenko, Byull. glavn. bot. Sada Leningr. 88: 98, 1973.
- Lovisolo & Conti, Annls Phytopath. 1: 367, 1969.
- Morales & Zettler, Fitopatologia Colombiana 6: 134, 1977.
- Tompkins & Severin, Hilgardia 20: 207, 1950.
- Tooyama, Ann. Phytopath. Soc. Japan 41: 504, 1975a.
- Tooyama, Ann. Phytopath. Soc. Japan 41: 506, 1975b.
- Van Hoof, Jversl. Inst. plziektenk. Onderz. Wageningen, 1971: 87, 1972.
- Volin & Zettler, HortScience 11: 459, 1976.
- Zettler, Foxe, Hartman, Edwardson & Christie, Phytopathology 60: 983, 1970.