Details of DPV and References
DPV NO: 199 August 1978
Family:
Genus:
Species: | Acronym:
Comovirus group
G. Bruening Department of Biochemistry and Biophysics, University of California, Davis, California 95616, USA
Contents
- Type Member
- Main Characteristics
- Members
- Geographical Distribution
- Transmission by Vectors
- Ecology and Control
- Relations with Cells and Tissues
- Properties of Particles
- Genome Properties
- Replication
- Satellite
- Defective-Interfering RNA
- Relationships
- Notes on Tentative Members
- Affinities with Other Groups
- Notes
- Acknowledgements
- Figures
- References
Type Member
Type member: cowpea mosaic virus, SB isolate.
Main Characteristics
Two types of ribonucleoprotein particle and, in some isolates, a protein particle, all c. 28 nm in diameter, all apparently with icosahedral symmetry, sedimenting at c. 118 S, 98 S and 58 S and containing c. 34, 25 and 0% single-stranded RNA, respectively. Two RNA species (with 3'-terminal polyadenylate sequences in most members examined), M. Wt c. 2.1 x 106 and 1.4 x 106, both necessary for infection. All three types of particle contain 60 molecules of each of two distinct coat polypeptides, M. Wt c. 22,000 and 42,000. Thermal inactivation point (10 min) 60-75°; survival in sap at 4-25° a few days to a few weeks; concentration in sap 10-1000 mg/l. Host range of an individual member typically only a few genera: usually symptoms are systemic mosaic or mottling and stunting, occasionally systemic wilting and collapse or necrotic ring symptoms or seed discoloration. Virus particles accumulate in the cytoplasm, sometimes in crystalline arrays. Readily transmissible by inoculation of sap; many members are transmitted by beetles and possibly other chewing insects, and some to progeny through seed.
Several members have been favoured objects of study by virologists; see reviews by Van Kammen (1972), Brown & Hull (1973), Jaspars (1974), and Bruening (1977).
Members
Members of the comovirus group and families of principal hosts
| Virus | Strains and synonyms | CMI/AAB Desc. or Ref. |
| Leguminosae (subclass Rosidae): | ||
| Bean curly dwarf mosaic virus (BCDMV) | Meiners et al., 1977 | |
| Bean pod mottle virus (BPMV) | type, J-10, Desmodium Virus | 108; Lee & Walters, 1970 | Bean rugose mosaic virus (BRMV) | type, Virus Ampollado del Frijol | 246; Gamez, 1972; Galvez et al., 1974 |
| Broad bean stain virus (BBSV) | 29 | |
| Cowpea severe mosaic virus (CPSMV) | Arkansas, Trinidad, Vu, Vs, El Salvador, DG | 47, 209 |
| Cowpea mosaic virus (yellow strain) (CPMV) | SB, Nigeria | 47, 197 |
| Echtes Ackerbohnemosaik-Virus (EAMV) | 20 | |
| Virus de la mosaique de la fève (MFV) | Devergne & Smith, 1964 | |
| Pea green mottle virus (PGMV) | F1 | Valenta et al., 1969 |
| Quail pea mosaic virus (QPMV) | 238; Moore, 1973 | |
| Red clover mottle virus (RCMV) | 74 | |
| Cruciferae (subclass Dilleniidae): | ||
| Radish mosaic virus (RaMV) | Neo-type, HZ, KV, B, DT4, S | 121; Plakolli & Štefanac, 1976 |
| Cucurbitaceae (subclass Dilleniidae): | ||
| Squash mosaic virus (SqMV) | 43 | |
| Solanaceae (subclass Asteridae): | ||
| Andean potato mottle virus (APMV) | 203; Fribourg et al., 1977 |
Geographical Distribution
Individual comoviruses tend to have a restricted distribution, in some instances probably dependent upon the distribution of vectors. As a group they have been reported in many tropical and temperate regions, especially where humidity is generally high. Host ranges of individual members are narrow. However, different members have natural hosts (Table 1) among four families representing three of the six subclasses (Cronquist, 1968) of dicotyledonous plants. Species of Chenopodium and Gomphrena (subclass Caryophyllidae) are local lesion hosts of many members. CPMV infects protoplasts from a range of cowpea cultivars, including some that are immune as intact plants (Beier et al., 1977). CPMV also infects tobacco protoplasts (Huber et al., 1977).
Transmission by Vectors
Most members are transmitted by leaf-feeding beetles. Transmission is a specific process dependent upon beetle species, virus strain and host (Fulton et al., 1975, 1980; Fulton & Scott, 1977). Beetles most commonly found to be efficient comovirus vectors are of the family Chrysomelidae (principally the genera Cerotoma and Diabrotica); vector species also occur in the families Curculionidae and Coccinellidae. Beetle vectors can acquire virus after feeding for 1 min and can retain it for days or weeks, or even several months if dormant (Fulton et al., 1975, 1980; Walters et al., 1972). The mechanisms of transmission are obscure; transmission is associated with feeding and the occurrence of virus in the haemolymph (Fulton & Scott, 1977). There is no evidence for replication in the vector. All the viruses in Table 1 except MFV, PGMV and APMV have beetle vectors. It is possible that some other chewing insects are comovirus vectors (Whitney & Gilmer, 1974).
Ecology and Control
Sources of virus causing initial infections in crops include seeds and perennial weed hosts, causing spread from the edges of the field, and perhaps overwintering beetles. Spread from an infected plant may occur in waves (Gibbs et al., 1968), implying possible involvement of beetles. Seed transmission has been reported for BBSV, CPSMV, CPMV, EAMV, MFV, and SqMV. Usually the frequency of seed transmission is less than 10% (e.g. Cockbain et al., 1976; Haque & Persad, 1975), but may be higher for SqMV. Partridge et al. (1974) have postulated that seed transmission is correlated with the capsid protein-bound carbohydrate.
Relations with Cells and Tissues
The comoviruses accumulate in highest concentrations in systemically infected leaves that were in bud or nor yet formed when the plant became infected. Somewhat lower concentrations are achieved in inoculated, expanded leaves and much lower concentrations in stems and other tissues. Some comoviruses invade developing seeds.
Properties of Particles
The two coat polypeptides, L and S, are present in equimolar amounts, apparently assembled in a T = 1 icosahedral structure containing 60 copies of each (Wu & Bruening, 1971; Crowther et al., 1974). Estimates of M. Wt vary from 37,000-49,000 (L) and 18,000-26,000 (S), but average about 42,000 and 22,000, respectively. In sodium dodecyl sulphate polyacrylamide gel electrophoresis the L proteins from several different comoviruses, and also the S proteins, comigrate. Carbohydrate is associated with the protein of CPMV (Partridge et al., 1974). Several members form c. 58 S ‘top’ components that lack RNA, suggesting a major contribution from protein-protein bonds in stabilizing the particles. Estimates of the capsid M. Wt are 4.5 x 106 for SqMV by sedimentation and diffusion (Mazzone et al., 1962) and 4.6 x 106 for CPMV by light-scattering (Geelen et al., 1972), implying larger M. Wt for the capsid proteins than the averages stated above. Members have two single-stranded RNA molecules, found both to be essential for infection where tested. The c. 118 S ‘bottom’ component contains one molecule of RNA-1 of M. Wt c. 2.1 x 106; the c. 98 S ‘middle’ component contains one molecule of RNA-2 of M. Wt c. 1.4 x 106. The components contain c. 34% and 25% RNA, respectively. Bottom component of some comoviruses forms two zones, BU and BL, in CsCl gradients during equilibrium centrifugation. The lower density of BU may reflect a higher content of polyamines, especially spermidine (Bruening, 1977).
At pH c. 9 several comoviruses can be resolved into two electrophoretic forms (e.g., Agrawal, 1964), each representing all of the centrifugal forms. The relative amount of the two forms varies with the time between inoculation and harvesting of the infected leaves. Conversion of one electrophoretic form to the other is associated with a reduction in the M. Wt of S polypeptide, presumably by proteolysis in vivo. In the pH range 7.5 to 8.2 CPMV top component was electrophoretically resolved from middle and bottom (reviewed by Bruening, 1977).
Genome Properties
Both RNA-1 and RNA-2, or the nucleoprotein components containing them, are required for infectivity; greater infectivity for mixtures of middle and bottom components than for the separated components has been observed for BPMV, BBSV, CPSMV, CPMV, EAMV, RCMV, RaMV (reviewed by Bruening, 1977) and APMV (Salazar & Harrison, 1978). If the viruses are sufficiently closely related, pseudo-recombinant isolates can be formed by inoculating a mixture of bottom component from one virus and middle component from another. Properties of the pseudo-recombinants show that RNA-2/middle component specifies some symptoms, the structure of S polypeptide (see, e.g., Gopo & Frist, 1977) and whether top component will be synthesized. RNA-1/bottom component specifies some symptoms and the kinetics of in vivo conversion of electrophoretic forms. Pseudo-recombinants do not form between CPSMV and CPMV or between the other pairs of distinct viruses that have been tested. Comovirus RNA is high in uridylate (30% or greater in most instances) and low in cytidylate. The RNA of CPMV and BPMV, and RNA-1 of RCMV (Oxelfelt, 1976), have polyadenylate sequences; these have been shown to be 3'-terminal in CPMV (El Manna & Bruening, 1973). The RNA species of CPMV have covalently bound protein at the 5'-termini (S. D. Daubert, G. Bruening and R. C. Najarian, unpublished data.) Molecular hybridization experiments suggest that RNA-1 and RNA-2 of CPMV have few extended nucleotide sequences in common (reviewed by Jaspars, 1974).
Replication
Most studies on replication have been made with CPMV. Coat (and presumably other) virus proteins are synthesized on cytoplasmic ribosomes because physiologically active concentrations of cycloheximide, but not of chloramphenicol, inhibit their synthesis (Owens & Bruening, 1975). Both RNA species are translated in vitro into polypeptides approaching in size the theoretical coding capacity (Davies et al., 1977); in vivo, such ‘polyproteins’ presumably are cleaved to form the functional proteins.
RNA complementary to particle RNA has been detected in infected tissue; the complementary RNA, particle antigen and probably RNA polymerase activity are all associated with cytoplasmic structures that tend to fractionate with, but can be resolved from, chloroplast fragments (De Zoeten et al., 1974; Zabel et al., 1974; Hibi et al., 1975). CPMV-induced RNA replicase has been partially purified (Zabel et al., 1976). Particles accumulate in crystalline aggregates (Langenberg & Schroeder, 1975).
Although Magyarosy et al. (1973) could find no functional change in chloroplasts from SqMV-infected tissue, the number of chloroplasts was reduced; Langenberg & Schroeder (1975) and De Zoeten et al. (1974) found lipid globules in chloroplasts of CPMV-infected tissue. Other changes in the host include a shift from carbohydrate to organic and amino acid synthesis (Magyarosy et al., 1973) and an increase in aspartate transcarbamylase activity (Niblett et al., 1974). CPMV and BPMV were found to be more, and more variably, sensitive to actinomycin D than most RNA viruses (Lockhart & Semancik, 1969).
Relationships
Serological relationships between members are generally distant. Where close relationships exist the assignments of isolates as the same virus or as different strains are difficult to make and unlikely to be clarified by further studies of symptoms, host range or vector specificity. Among the serological relationships shown in Fig. 1, those between BBSV, MFV and PGMV are closest and they should perhaps be considered to be strains of one virus; BBSV and MFV both invade developing seeds and cause discoloration (Desc. 29; Devergne & Cousin, 1966). QPMV and BCDMV are reported to be closely serologically related (Meiners et al., 1977). Assignment of RaMV isolates to either the neotype (isolates from USA and Japan) or European type on the basis of serology, host range and pseudo-recombinant formation has been suggested (Plakolli & Stefanac, 1976).
The distinction between CPSMV and CPMV is based upon symptoms, serology, and, generally, a lack of top component, a higher middle to bottom component ratio, a lower specific infectivity and a wider host range for CPSMV than for CPMV (Agrawal, 1964; Swaans & Van Kammen, 1973; Beier et al., 1977). Most CPMV isolates are from the Old World, whereas most CPSMV isolates are from the New World (Fulton et al., 1975). A few exceptions have been reported (Agrawal, 1964; Williams, 1975; McLaughlin et al., 1977).
Affinities with Other Groups
Broad bean wilt virus (BBWV) is very similar to the comoviruses. The L and S coat proteins of BBWV co-migrated with those of CPMV during electrophoresis in polyacrylamide gels, as did the respective RNA-1 and RNA-2. BBWV forms a top component, and evidence was obtained of a requirement for both middle and bottom component in initiating infections. The RNA has a high uridylate to cytidylate ratio (Doel, 1975). However, unlike the comoviruses, BBWV reaches only moderate concentrations in infected plants, is transmitted by aphids and has a wide host range. Comoviruses also resemble, though less closely, the nepovirus group. Strawberry latent ringspot virus is the most similar to the comoviruses because, unlike other nepoviruses, it has two coat proteins close to the sizes of the comovirus proteins. An even more distant resemblance is seen to the animal picornaviruses (e.g. poliovirus), which exhibit 3'-terminal polyadenylate, a 5'-linked protein, more than one coat protein, and expression of the virus genome via polyproteins.
Figures
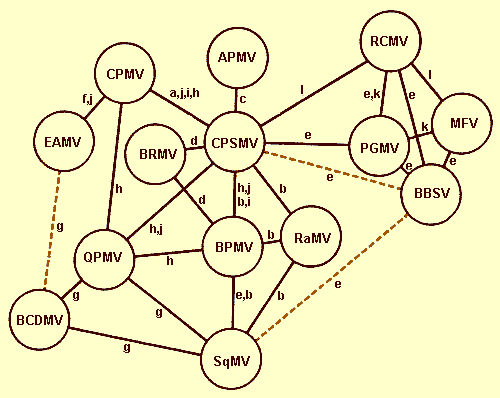
Diagram of serological relationships between comoviruses. Solid lines represent
serological relationships in which the heterologous titre was 1/64th or
more of the homologous
titre or in which an immunodiffusion pattern was judged to show a precipitin line whose
intensity for the heterologous reaction was not much less than that for the homologous
reaction. Dashed lines represent serological relationships that meet these citeria barely
or questionably
References:
(a) Agrawal & Maat, 1964;
(b) Campbell, 1964;
(c) Fribourg et al., 1977;
(d) Gamez, 1972;
(e) Gibbs et al., 1968;
(f) Jones & Barker, 1976;
(g) Meiners et al., 1977;
(h) Moore, 1973;
(i) Shepherd, 1963;
(j) D.J. Siler, H.A. Scott & G. Bruening, unpublished observations;
(k) Valenta et al., 1969;
Valenta & Gressnerová, 1966;
(l) Desc. 74.
References list for DPV: Comovirus group (199)
- Agrawal, Meded. LandbHoogesch. Wageningen 64: 1, 1964.
- Agrawal & Maat, Nature, Lond. 202: 674, 1964.
- Beier, Siler, Russell & Bruening, Phytopathology 67: 917, 1977.
- Brown & Hull, J. gen. Virol. 20 Suppl.: 43, 1973.
- Bruening, in Comprehensive Virology, Vol. 11, p. 55. ed. H.Fraenkel-Conrat & R. R. Wagner, New York: Plenum, 1977.
- Campbell, Phytopathology 54: 1418, 1964.
- Cockbain, Bowen & Vorra-Urai, Ann. appl. Biol. 84: 321, 1976.
- Cronquist, The Evolution and Classification of Flowering Plants. New York: Houghton Mifflin, 1968.
- Crowther, Geelen & Mellema, Virology 57: 20, 1974.
- Davies, Aalbers, Stuik & Van Kammen, FEBS Lett. 77: 265, 1977.
- Devergne & Cousin, Annls Épiphyt. numéro hors série: 147, 1966.
- Devergne & Smith, Études de Virologie appliquée 5: 7, 1964.
- De Zoeten, Assink & Van Kammen, Virology 59: 341, 1974.
- Doel, J. gen. Virol. 26: 95, 1975.
- El Manna & Bruening, Virology 56: 198, 1973.
- Fribourg, Jones & Koenig, Phytopathology 67: 969, 1977.
- Fulton & Scott, Fitopathol. Brasileira 2: 9, 1977.
- Fulton, Scott & Gamez, in Tropical Diseases of Legumes. p. 123. ed. J. Bird & K. Maramorosch, New York: Academic Press, 1975.
- Fulton, Scott & Gamez, in Vectors of Plant Disease Agents, p. 115, ed. K. Maramorosch & K. F. Harris, New York: Academic Press, 1980.
- Galvez, Cardenas & Diaz, Proc. Am. Phytopath. Soc. 1: 122, 1974.
- Gamez, Turrialba 22: 249, 1972.
- Geelen, Van Kammen & Verduin, Virology 49: 205, 1972.
- Gibbs, Giussani-Belli & Smith, Ann. appl. Biol. 61: 99, 1968.
- Gopo & Frist, Virology 79: 259, 1977.
- Haque & Persad, in Tropical Diseases of Legumes, p. 119, ed. J.Bird & K. Maramorosch, New York: Academic Press, 1975.
- Hibi, Rezelman & Van Kammen, Virology 64: 308, 1975.
- Huber, Rezelman, Hibi & Van Kammen, J. gen. Virol. 34: 315, 1977.
- Jaspars, Adv. Virus Res. 19: 37, 1974.
- Jones & Barker, Ann. appl. Biol. 83: 231, 1976.
- Langenberg & Schroeder, J. Ultrastruct. Res. 51: 166, 1975.
- Lee & Walters, Phytopathology 60: 585, 1970.
- Lockhart & Semancik, Virology 39: 362, 1969.
- McLaughlin, Thongmeearkom, Milbrath & Goodman, Phytopathology 67: 844, 1977.
- Magyarosy, Buchanan & Schumann, Virology 55: 426, 1973.
- Mazzone, Incardona & Kaesberg, Biochim. biophys. Acta 55: 164, 1962.
- Meiners, Waterworth, Lawson & Smith, Phytopathology 67: 163, 1977.
- Moore, Pl. Dis. Reptr 57: 311, 1973.
- Niblett, Johnson & Lee, Physiol. Pl. Path. 4: 63, 1974.
- Owens & Bruening, Virology 64: 520, 1975.
- Oxelfelt, Virology 74: 73, 1976.
- Partridge, Shannon, Gumpf & Colbaugh, Nature, Lond. 247: 391, 1974.
- Plakolli & Stefanac, Phytopath. Z. 87: 114, 1976.
- Salazar & Harrison, J. gen. Virol. 39: 171, 1978.
- Shepherd, Phytopathology 53: 865, 1963.
- Swaans & Van Kammen, Neth. J. Pl. Path. 79: 257, 1973.
- Valenta & Gressnerová, Acta Virol. 10: 182, 1966.
- Valenta, Gressnerová, Marcinka & Nermut, Acta Virol. 13: 422, 1969.
- Van Kammen, A. Rev. Phytopath. 10: 125, 1972.
- Walters, Lee & Jackson, Phytopathology 62: 808, 1972.
- Whitney & Gilmer, Ann. appl. Biol. 77: 17, 1974.
- Williams, in Tropical Diseases of Legumes, p. 139, ed. J. Bird & K. Maramorosch, New York: Academic Press, 1975.
- Wu & Bruening, Virology 46: 596, 1971.
- Zabel, Weenen-Swaans & Van Kammen, J. Virol. 14: 1049, 1974.
- Zabel, Jongen-Neven & Van Kammen, J. Virol. 17: 679, 1976.