Details of DPV and References
DPV NO: 202 July 1979
Family: Unallocated ssRNA+ viruses
Genus: Unassigned ssRNA+ virus
Species: Lilac chlorotic leafspot virus | Acronym: LiCLV
Lilac chlorotic leafspot virus
A. A. Brunt Glasshouse Crops Research Institute, Littlehampton, Sussex, England
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
Described by
Brunt (1978) and
Brunt & Stace-Smith (1978).
Synonym
- Lilac virus St. 30 (Rev. Pl. Path. 56: 2092)
-
A virus with fragile filamentous particles c. 1540 x 12.5 nm. The particles occur in mesophyll and phloem cells, usually in inclusions also containing cytoplasm with extensively proliferated endoplasmic reticulum. The virus is transmissible manually to 21 species from eight families, but its vector is unknown. Found in Europe.
Main Diseases
Only known from lilac (Syringa vulgaris). Associated with chlorotic leaf-spotting symptoms (Fig. 1) in an unnamed cultivar in S.E. England, but isolated in the Netherlands from plants that were either symptomless or had only faintly chlorotic leaves (Van der Meer, 1976).
Geographical Distribution
Reported only from England and the Netherlands, but it possibly occurs elsewhere in Europe (Van der Meer, 1976) and the USA (Beale & Beale, 1952).
Host Range and Symptomatology
Transmitted easily by mechanical inoculation to 21 of 52 species from 8 of 20 families (Brunt, 1978).
-
Diagnostic species
- Beta vulgaris
cv. Detroit (red beet). Irregular necrotic local lesions after c. 15 days, soon followed by faint systemic leaf chlorosis. B. vulgaris cv. Monobush (sugarbeet) is not susceptible. -
Chenopodium capitatum, C. murale. Few necrotic local lesions after 10-15 days;
no systemic infection.
- C. quinoa. Irregular necrotic local lesions after 12-15 days (Fig. 2), soon followed by faint systemic leaf chlorosis.
- Nicotiana clevelandii. Faint systemic leaf chlorosis after 15-25 days.
- Phaseolus vulgaris (French bean). Inoculated leaves infected symptomlessly; systemically infected leaves develop chlorotic leaf-spots which may coalesce to produce almost complete chlorosis (Fig. 3).
- Trifolium incarnatum (crimson clover). Transient chlorotic vein-banding patterns on young systemically infected leaves, and permanent chlorotic spots on older leaves.
- C. quinoa. Irregular necrotic local lesions after 12-15 days (Fig. 2), soon followed by faint systemic leaf chlorosis.
-
Propagation species
- Chenopodium quinoa
is a good source of virus for purification and for maintaining cultures.Assay species
- Chenopodium quinoa
can be used as an assay host, but the production of local lesions is erratic.
Strains
No reports.
Transmission by Vectors
Not transmitted by the aphids Aphis gossypii, Macrosiphum sanborni, Myzus ligustri or Myzus persicae (Brunt, 1978), although the affinities of the virus suggest that it may be aphid-borne in the semi-persistent manner.
Transmission through Seed
Not seed-borne in Syringa vulgaris, Chenopodium quinoa, Nicotiana clevelandii x N. glutinosa hybrid, or Phaseolus vulgaris (Brunt, 1978).
Serology
The virus is a good immunogen. Antiserum obtained from a rabbit injected intradermally (three injections at monthly intervals, each of c. 0.06 mg of virus emulsified in 0.3 ml Freund’s complete adjuvant) reacted with homologous virus in tube precipitin and microprecipitin tests to produce flagellar precipitates; the antiserum titre was 1/1024.
Relationships
Lilac chlorotic leafspot virus and lilac virus St. 30 (Van der Meer, 1976) are serologically indistinguishable (D. Z. Maat & A. A. Brunt, unpublished information). The virus has very flexuous filamentous particles similar in size, morphology, physical properties and distribution in vivo to those of closteroviruses but it showed no serological relationship to three definitive members of the group (beet yellow stunt, beet yellows and carnation necrotic fleck viruses), nor to three other viruses with very flexuous particles (apple chlorotic leafspot, apple stem grooving or potato T) (Brunt, 1978). Also, it failed to infect the major hosts of the above viruses, or of three other closteroviruses (citrus tristeza, wheat yellow leaf or festuca necrosis). Lilac chlorotic leafspot virus is thus probably a newly recognized member of the closterovirus group. However, unlike some closteroviruses, it is readily transmissible by manual inoculation, is moderately stable in vitro and has not been transmitted in the semi-persistent manner by aphids.
Stability in Sap
In Chenopodium quinoa sap, infectivity is lost after 10 min at 60-65°C, 8-16 days at 20°C or 25-30 weeks at 2°C, or after dilution between 10-3 and 10-4. Sap from Syringa vulgaris has a maximum infective dilution of only 10-1 and is often non-infective after 1 h at 2°C; infectivity can be preserved for up to 20 h in the presence of a reducing agent (0.05 M thioglycollate or 2-mercaptoethanol) (Brunt, 1978).
Purification
Purified preparations of the virus are best obtained from inoculated and systemically infected leaves of Chenopodium quinoa seedlings 12-21 days after inoculation. The following procedure yields up to 18 mg per kg leaf tissue (Brunt, 1978). Homogenize infected leaves (1 g/2 ml) in 0.07 M phosphate buffer (pH 7.6) containing 0.14 M thioglycollate, and clarify by adding n-butanol to 8% (v/v). Subject the extracts to one cycle of differential centrifugation (15 min at 12,000 g; 75 min at 65,000 g) and resuspend the final pellets in 0.03 M neutral phosphate buffer (1 ml/40-50 g leaf material). Further purify the virus by permeation chromatography on columns of controlled-pore glass beads (700 Å, 120-200 mesh) as described by Barton (1977), and concentrate the virus from appropriate eluate fractions by centrifugation at 65,000 g for 75 min.
Properties of Particles
Sedimentation coefficient (s°20,w): 96 S. No accessory particles are found by analytical ultracentrifugation.
Buoyant density in CsCl: 1.302 g/cm3.
Amax. (260)/Amin. (242): 1.69; A260/A280: 1.69. The high A260/A280 ratio, like that of beet yellows virus, is presumably due to a low concentration of aromatic amino acids in the protein.
Particle Structure
Particles are fragile flexuous filaments. Particles that are apparently intact measure 12.5 x 1540 nm and have a helical substructure with a pitch of c. 3.7 nm (Fig. 6).
Particle Composition
Nucleic acid: Buoyant density estimations suggest that the virus contains c. 4.5-5.0% nucleic acid.
Protein: Electrophoresis in polyacrylamide/SDS gels indicates that the protein subunit is a polypeptide of M. Wt 27 x 103.
Relations with Cells and Tissues
In Chenopodium quinoa and Phaseolus vulgaris the filamentous virus particles occur in mesophyll and phloem parenchyma cells, but apparently not in sieve tubes or other cell types (Brunt & Stace-Smith, 1978). In immature cells the particles are randomly distributed in central cytoplasmic masses or in peripheral cytoplasm. In fully differentiated cells the particles occur only within ovoid inclusions; these, usually one per cell, are often adjacent to the nucleus. Inclusions are up to 8 µm in diameter in mature mesophyll cells and 3-4 µm in diameter in phloem parenchyma cells, and in both cell types consist of a cytoplasmic matrix containing numerous randomly distributed filamentous virus particles and extensively proliferated and dilated endoplasmic reticulum (Fig. 4, Fig. 5). The inclusions are not membrane-bounded but in fully differentiated cells they often extend in peripheral cytoplasm from the tonoplast to the plasmalemma.
Notes
Eight other sap-transmissible viruses are known to infect Syringa vulgaris (Waterworth, 1972; Van der Meer, 1976; Novák & Lanzová, 1977; Brunt, 1978). Six of these (arabis mosaic, cherry leaf roll, elm mottle, lilac ring mottle, strawberry latent ringspot and tomato bushy stunt) have isometric particles 25-30 nm in diameter, one (tobacco rattle virus) has rod-shaped particles, and the other (lilac mottle virus) has filamentous particles c. 650 nm long. These viruses are thus morphologically different from lilac chlorotic leafspot virus, and are also readily distinguishable from it by their different host ranges, serological affinities and physico-chemical properties.
The virus apparently occurs only rarely in S.E. Britain. The availability of simple methods for its detection and identification suggests that it could be eradicated by selecting only known virus-free material for propagation.
Acknowledgements
Micrographs 4 and 5 by courtesy of Canada Agriculture Research Station, Vancouver, Canada.
Figures

Systemic chlorotic leaf-spotting in naturally-infected Syringa vulgaris.

Irregular necrotic lesions in inoculated leaf of Chenopodium quinoa.
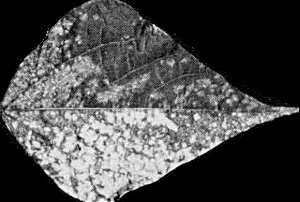
Systemic leaf chlorosis in Phaseolus vulgaris cv. The Prince.

Sections through intracellular inclusions at lower magnification, showing numerous filamentous virus particles distributed throughout a cytoplasmic matrix containing proliferated and dilated endoplasmic reticulum. Bars represent 500 nm.
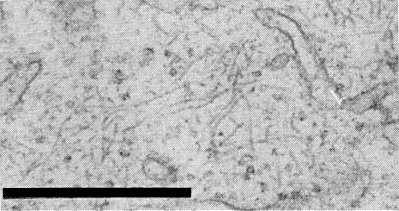
Sections through intracellular inclusions at higher magnification, showing numerous filamentous virus particles distributed throughout a cytoplasmic matrix containing proliferated and dilated endoplasmic reticulum. Bars represent 500 nm.

Intact particle mounted in neutral potassium phosphotungstate (bar represents 500 nm). Inset, particle fragments mounted in uranyl acetate showing helical substructure (bar represents 25 nm).
References list for DPV: Lilac chlorotic leafspot virus (202)
- Barton, J. gen. Virol. 35: 77, 1977.
- Beale & Beale, Contr. Boyce Thompson Inst. Pl. Res. 17: 1, 1952.
- Brunt, Ann. appl. Biol. 88: 383, 1978.
- Brunt & Stace-Smith, J. gen. Virol. 39: 63, 1978.
- Novák & Lanzová, Biol. Plant. 19: 264, 1977.
- Van der Meer, Acta Hort. 59: 105, 1976.
- Waterworth, Pl. Dis. Reptr 56: 923, 1972.