Details of DPV and References
DPV NO: 204 July 1979
Family: Unallocated ssRNA+ viruses
Genus: Sobemovirus
Species: Blueberry shoestring virus | Acronym: BSSV
Blueberry shoestring virus
D. C. Ramsdell Department of Botany and Plant Pathology, Michigan State University, East Lansing, Michigan, USA
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
Described by Varney (1957); Lesney & Ramsdell (1976); Lesney, Ramsdell & Sun (1978).
An RNA-containing virus with isometric particles about 27 nm in diameter. Found only in Vaccinium corymbosum and V. angustifolium. The virus is transmissible by manual inoculation of purified preparations to blueberry seedlings or rooted soft-wood cuttings. Not manually transmissible to herbaceous plants. Transmitted by the aphid Masonaphis pepperi. Found in USA and Canada.
Main Diseases
Causes ‘shoestring’ disease of highbush blueberry (Vaccinium corymbosum). There is one report of infection in lowbush blueberry (V. angustifolium) (Lockhart & Hall, 1962). Symptoms consist of elongated (0.2 x 1.2 cm) reddish streaks on current year and one-year-old stems, especially on the side exposed to the sun (Fig. 1). As the growing season progresses the stem lesions may disappear. Occasionally, flower ‘breaking’ occurs in the form of longitudinal pink streaks on the petals. Affected leaves are narrow and strap-like (‘shoestring’ symptom; Fig. 2) or curled or crescent-shaped. Some bushes show extensive leaf symptoms but others exhibit only an occasional cluster of symptom-bearing leaves. A few leaves may show red vein-banding or reddish streaking along the mid-rib and, on occasion, oak-leaf patterns. Immature berries on infected plants may develop a premature reddish-purple cast on the surface exposed to light. Yield of infected bushes is much decreased. The disease spreads along the row from bush to bush (Lesney et al., 1978). Healthy bushes planted in a field with disease show symptoms after about 4 years.
Geographical Distribution
New Jersey, Michigan (Stretch & Hilborn, 1970) and Washington States, USA (P. R. Bristow & D. C. Ramsdell, unpublished data) and Nova Scotia, Canada (Lockhart & Hall, 1962).
Host Range and Symptomatology
Blueberries are the only hosts known. Shoestring disease has been observed in the highbush blueberry cultivars Burlington, Coville, Earliblue, Jersey, June, Rancocas, Rubel and Weymouth. The virus was transmitted by manual inoculation of virus purified from infected blossoms to healthy seedlings and to rooted soft-wood cuttings of blueberry cv. Jersey, which showed symptoms within 5 or 6 months, but was not transmitted to a wide range of herbaceous species (Lesney & Ramsdell, 1976; Lesney et al., 1978). Blueberries infected by chip-budding (Varney, 1957) show symptoms within 1 year. Under field conditions, the highbush cultivars Bluecrop and Atlantic do nor become infected but when seedlings of these cultivars were manually inoculated with purified blueberry shoestring virus a low percentage developed symptoms within 5 or 6 months (D. C. Ramsdell, unpublished data).
Strains
None reported.
Transmission by Vectors
Transmitted by the blueberry aphid Masonaphis pepperi, after a 2 min acquisition feed and a 100 h inoculation feed, to two out of five rooted soft-wood blueberry cuttings (D. C. Ramsdell, unpublished data).
Transmission through Seed
Apparently none in V. corymbosum (Lesney et al., 1978).
Serology
The virus is a good immunogen. A rabbit given three intramuscular injections at 2-week intervals of approximately 0.1 mg virus emulsified 1:1 with Freund’s complete adjuvant, yielded antiserum with a titre of 1/256 in agar gel double diffusion tests (Lesney & Ramsdell, 1976; Lesney et al., 1978). The virus forms a single precipitin band in agar gel-diffusion tests. Virus is readily detected in blueberry tissue using enzyme-linked immunosorbent assay (ELISA) (D. C. Ramsdell, unpublished data).
Relationships
The virus gave no reaction when tested in agar double diffusion tests against antisera to thirty-three viruses with isometric particles, including members of the bromovirus, comovirus, cucumovirus, ilarvirus, nepovirus, tombusvirus and tymovirus groups and several unclassified viruses (Lesney et al., 1978; D. C. Ramsdell, unpublished data). Antiserum to blueberry shoestring virus did not react with any of twenty-four of these viruses.
Stability in Sap
No information.
Purification
The best method is as follows (Ramsdell, 1979). All stages of virus purification should be done at 4°C; at higher temperatures the virus aggregates (reversible at 4°C after a few hours). Disrupt infected blossom tissue (1:3 w/v) in a blender with 0.1 M phosphate buffer, pH 7.0, containing 0.1 M 2-mercaptoethanol and 0.005 M sodium thioglycollate. Add Triton X-100 to 8.0% (v/v) and stir for 2 h. Squeeze through cheesecloth. Add chloroform and n-butanol (each to 5.0% (v/v)), and stir thoroughly for 15 min. Centrifuge at low speed for 15 min, retaining the aqueous supernatant fluid. Add polyethylene glycol (M. Wt 6000) to 8.0% (w/v) and NaCl to 0.1 M. Stir and let stand 30 min, then centrifuge at low speed for 30 min. Resuspend the pellet in 1/10 of the starting volume with 0.05 M phosphate buffer, pH 7.0, containing 0.001 M dithiothreitol. Let stand for several hours. Centrifuge at low speed for 20 min, retaining the supernatant fluid. Centrifuge at 108,000 g for 2 h, and resuspend pellet in 0.05 M phosphate buffer, pH 7.0, containing 0.001 M dithiothreitol. Purify further by centrifugation in a 5-30% sucrose gradient in a Beckman SW41 rotor for 90 min at 38,000 rev./min or in a SW25.1 rotor for 3 h at 23,000 rev./min. Collect the single virus-containing zone from the gradient, dilute it three or four-fold with buffer and centrifuge at 137,000 g for 2 h to pellet the virus. Resuspend pellet in 0.05 M phosphate buffer at pH 7.0. Store with 0.1% chlorobutanol as a preservative at 4°C. Average yield is about 5 mg/kg blossoms.
Properties of Particles
Sedimentation coefficient: (s20,w) about 120 S at infinite dilution.
M. Wt: approximately 6.9 x 106 (by calculation, assuming that the particle contains 180 protein subunits).
Isoelectric point: between pH 4.6 and 4.7.
Electrophoretic mobility: 0.42 x 10-5 cm2 sec-1 volt-1 at pH 5.0 in 0.02 M Na2HPO4, 0.02 M Tris buffer, titrated with citric acid. Determinations were made in 0.7% agarose slab gels. The virus migrates as a single electrophoreric component.
Absorbance at 260 nm (1 mg/ml, 1 cm light path): 5.2.
A260/A280 = 1.47 (not corrected for light-scattering).
Buoyant density in CsCl: 1.392 g/cm3.
Buoyant density in Cs2SO4: 1.273 and 1.355 g/cm3 (forms two bands).
Particle Structure
Isometric particles 27 nm in diameter (Fig. 3) (Ramsdell, 1979). No obvious surface structure is revealed in phosphotungstate negative stain.
Particle Composition
Nucleic acid: RNA, M. Wt about 1.45 x 106, estimated by electrophoresis in 2.6% polyacrylamide gels and comprising about 20% (by phosphorus determination) of the particle weight. The thermal denaturation profile indicates that the RNA is single-stranded (hyperchromicity is 13.6% from 50 to 99.5°C) and has a Tm of 63.6°C in 0.1 M NaCl.
Protein: Subunits have M. Wt about 30,000, estimated by electrophoresis in 5.0% polyacrylamide/SDS gels, and comprise about 80% of the particle weight.
Relations with Cells and Tissues
(Hartmann, Bath & Hooper, 1973). In diseased blueberry leaf tissues, virus-like particles (VLP) were found in epidermal cells in small crystalline arrays and as individual particles. In palisade parenchyma cells, VLP were found in small cytoplasmic vacuoles and were also associated with chloroplast membranes (Fig. 4). Elongated tubules or aggregates of rod-like materials were found in diseased palisade cells (Fig. 5), both in the nuclei and in the cytoplasm. In leaves, VLP occurred in xylem parenchyma, confined to small clear areas in the periphery of the cells but not in mature xylem vessels or in leaf phloem tissue. In root tissues, VLP were present in varying amounts in almost all xylem parenchyma cells examined, usually in greater concentrations than in leaf xylem parenchyma (Fig. 6). VLP have not been found in the roots in other types of cell. No VLP were found in plasmodesmata.
Notes
Although serologically unrelated to previously described viruses, blueberry shoestring virus has many properties in common with those comprising the proposed southern bean mosaic virus group (Hull, 1977). The virus forms one band in CsCl, but forms two or more bands in Cs2SO4 gradients, is stabilized by divalent cations (Ramsdell, 1979) and forms crystalline arrays in the cytoplasm of its host cells (Hartmann et al., 1973). However, whereas some of the viruses of the southern bean mosaic group are transmitted by beetles, preliminary experiments suggest that blueberry shoestring virus is transmitted by aphids. The pattern of spread of the virus in the field suggests that it is spread by apterae.
Acknowledgements
Figs 4, 5, 6 Courtesy of J. X. Hartmann, Dept. of Biology, Florida Atlantic University, Boca Raton, Florida.
Figures

Reddish, elongated stem lesions (arrow) on infected highbush blueberry cv. Jersey.

Narrow strap-like leaves on infected highbush blueberry cv. Jersey.
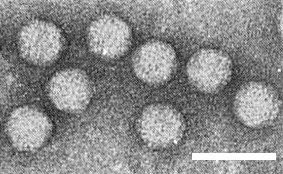
Virus particles in 2.0% uranyl acetate. Bar represents 50 nm.

Virus-like particles (P) associated with chloroplasts in palisade parenchyma cell from infected blueberry leaf. Tubules (T) are shown in cross-section. Bar represents 250 nm.
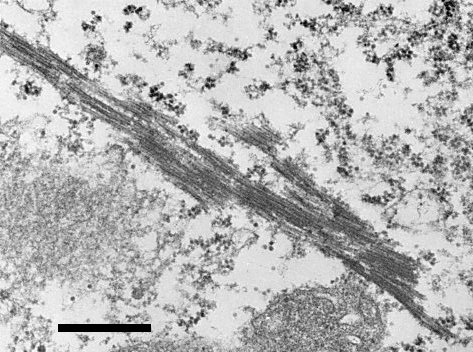
Longitudinal section of tubules (rod-like) in palisade cell of an infected blueberry leaf. Bar represents 500 nm.

Xylem parenchyma fibre cell from the root of an infected blueberry bush showing large mass of virus-like particles. Bar represents 500 nm.
References list for DPV: Blueberry shoestring virus (204)
- Hartmann, Bath & Hooper, Phytopathology 63: 432, 1973.
- Hull, Virology 36: 289, 1977.
- Lesney & Ramsdell, Acta Hort. 66: 105, 1976.
- Lesney, Ramsdell & Sun, Phytopathology 68: 295, 1978.
- Lockhart & Hall, Can. J. Bot. 40: 1561, 1962.
- Ramsdell, Phytopathology 69: 1087, 1979.
- Stretch & Hilborn, in Virus Diseases of Small Fruits and Grapevines, p. 186, University of California Press, 1970.
- Varney, Phytopathology 47: 307, 1957.