Details of DPV and References
DPV NO: 205 July 1979
Family: Rhabdoviridae
Genus: Nucleorhabdovirus
Species: Sonchus yellow net virus | Acronym: SYNV
Sonchus yellow net virus
A. O. Jackson Botany and Plant Pathology Department, Purdue University, West Lafayette, IN 47907, USA
S. R. Christie Plant Virus Laboratory, Plant Pathology Department, University of Florida, Gainesville, Florida 32611, USA
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
Described by Christie, Christie & Edwardson (1974).
Synonym
- Sowthistle yellow net virus (Rev. Pl. Path. 55: 3966)
-
A virus with bacilliform particles which contain protein, RNA, carbohydrate and probably lipid. The virus is transmissible by aphids and by mechanical inoculation of plant extracts containing a reducing agent. Occurs in USA.
Main Diseases
Occurs naturally in certain weeds of the family Compositae including Bidens pilosa, Sonchus oleraceus (sowthistle) and Senecio glaubellus. All hosts found infected in nature have also been infected with bidens mottle virus. These plants showed various degrees of stunting together with veinal necrosis and yellowing of leaves. Sonchus yellow net virus alone causes severe stunting and mosaic symptoms in experimentally infected lettuce (Lactuca sativa).
Geographical Distribution
Central and south Florida, USA.
Host Range and Symptomatology
Known to infect six species of Compositae, four Nicotiana spp. (Solanaceae), and also Chenopodium quinoa (Chenopodiaceae).
-
Diagnostic species
- Chenopodium quinoa.
Tan-coloured local lesions about 1 mm in diameter develop in mechanically inoculated leaves in 8-12 days (Fig. 2). The virus is recoverable from single lesions, but only up to 1 or 2 days after they become visible. Young uninoculated leaves usually develop a mild systemic mottle in 3-4 weeks. - Nicotiana hybrid (N. clevelandii x N. glutinosa)
(Christie, 1969).
Shows faint chlorotic local lesions. Lesions are less discrete than those in
S. oleraceus but the Nicotiana hybrid is more susceptible to infection
by inoculation with sap (Christie et al., 1974).
Systemic chlorotic vein-netting (Fig. 3)
is accompanied by thickening and downward cupping of leaves. Similar symptoms
are induced in N. clevelandii and N. benthamiana.
- Sonchus oleraceus. Discrete chlorotic local lesions followed by systemic chlorotic vein-clearing (Fig. 4).
-
Propagation species
- Cultures have been maintained without transfer in the Nicotiana hybrid for periods exceeding 6 months. This hybrid and N. benthamiana can be used as sources for virus purification (Jackson & Christie, 1977; Jackson, 1978).
-
Assay species
- Chenopodium quinoa
is useful as a local lesion host (Jackson, 1978) but lesion production is extremely dependent on age of the plant, position of the leaf and on duration and intensity of light. The Nicotiana hybrid is a susceptible host for systemic assay (Christie et al., 1974).
Strains
None reported.
Transmission by Vectors
Transmitted by Aphis coreopsidis, which colonizes Bidens pilosa. Persistence of the virus in aphids has not been studied critically but the virus is believed to multiply in the vector because bacilliform particles are found in the cells of viruliferous aphids. Not transmitted by either Hyperomyzus lactucae or a Dactanotus sp., both of which colonize Sonchus oleraceus (Christie et al., 1974).
Transmission through Seed
Not detected in Sonchus oleraceus (S. R. Christie, unpublished data).
Serology
Rabbits immunized with purified virus yielded antisera that reacted in gel double diffusion tests with sonchus yellow net virus disrupted with 0.5% sodium dodecyl sulphate (Jackson & Christie, 1977). The virus appears to be a poor immunogen because the highest antiserum titre observed in these tests was 1/32.
Relationships
Particle morphology places sonchus yellow net virus in the plant rhabdovirus group (Christie et al., 1974; Fenner, 1976). The virus resembles sowthistle yellow vein virus more closely than other plant rhabdoviruses in host range and in the size, structure and protein composition of its particles (Jackson & Christie, 1977; Jackson, 1978). However, the two viruses are serologically distinct, differ in mechanical transmissibility and have different aphid vectors. Sonchus yellow net virus fails to react with antisera to broccoli necrotic yellows, lettuce necrotic yellows or potato yellow dwarf viruses. Sonchus yellow net and sowthistle yellow vein viruses resemble eggplant mottled dwarf and potato yellow dwarf viruses in having two membrane matrix proteins whereas broccoli necrotic yellows virus, sonchus virus (Vega et al., 1976) and lettuce necrotic yellows virus reportedly have only one matrix protein (Peters, Toriyama & Terlouw, 1978).
Stability in Sap
The infectivity of sonchus yellow net virus is quickly lost unless leaf tissue is ground in the presence of a reducing agent such as 0.5% sodium sulphite (Christie et al., 1974; Jackson & Christie, 1977). In the presence of sodium sulphite, infectivity of leaf extracts is maintained for less than 8 h at 20°C but for more than 48 h at 4°C. Dilution end-point is 10-3 to 10-4.
Purification
Jackson & Christie (1977). Harvest systemically infected leaves of the Nicotiana hybrid or N. benthamiana 9-12 days after inoculation, store for several days at 4°C or extract immediately. Blend leaf tissue for 1 min with 2 vol cold 0.1 M Tris-HCl, pH 8.4, containing 0.01 M Mg acetate, 0.04 M Na2SO3 and 0.001 M MgCl2. Remove the brei by low speed centrifugation and centrifuge the supernatant fluid in discontinuous gradients formed from layers of 300 and 600 mg/ml sucrose. Clarify the fraction between the 300 and 600 mg/ml layers by filtering through a Celite pad and sediment the virus by centrifuging at 50,000 g for 30 min. Adjust the buffer used for extraction to pH 7.5 and resuspend the small tan-coloured pellets in 1 ml of buffer/100 g of tissue. Purify the virus further by centrifugation into rate zonal and quasi-equilibrium sucrose gradients. The final preparation has a milky appearance and is essentially free of host membranes or ribosomes. Yield is usually 2-5 A260 units/100 g tissue but is very dependent on age of the plants when inoculated, light and temperature in the greenhouse, and time after inoculation. A particularly critical variable in the purification procedure is the thickness of the Celite pad used for filtration. Celite pads thicker than 7.5 mm seriously diminish yield of virus, but pads less than 2.5 mm result in filtrates contaminated with chloroplast fragments.
Properties of Particles
Particles sediment as a homogeneous band with a sedimentation coefficient (s20,w) of approximately 1050 S.
Buoyant density in sucrose is 1.18 g/cm3.
M. Wt of the virus particle, estimated from size and density, is about 9 x 108 (Jackson & Christie, 1977).
Particle Structure
Bacilliform particles (Fig. 1) can be seen in leaf dip preparations from infected plants. Particles in purified preparations (Fig. 5) measure 94 x 248 nm after fixation in glutaraldehyde and negative staining (Jackson & Christie, 1977). The particles have internal cross-striations with a periodicity of 4.1 nm and an outer envelope through which 6 nm-long surface projections protrude.
Particle Composition
Nucleic acid: RNA, single-stranded, with M. Wt 4.4 x 106 as estimated by polyacrylamide gel electrophoresis, and a sedimentation rate (s20, w) of 44 S (Jackson & Christie, 1977). The RNA content has not been determined but is thought to be less than 1%. The RNA in the particles is the negative strand [i.e. complementary to messenger RNA isolated from polyribosomes (Milner & Jackson, 1979)].
Protein: Four major polypeptides with M. Wt about 77, 64, 45 and 39 kilodaltons are resolved by polyacrylamide gel electrophoresis of disrupted virus particles (Jackson & Christie, 1977). These have properties of a surface glycoprotein (G), a nucleic acid-associated protein (N) and two membrane matrix proteins (M1 and M2) respectively (Jackson, 1978). In addition, three minor proteins of unknown origin are found consistently in purified preparations. The proteins of sonchus yellow net virus and potato yellow dwarf virus (Black, 1970) are similar in number but differ in their electrophoretic mobility (Fig. 6).
Carbohydrate: The G protein (77 kilodaltons) reacts with stains for carbohydrate but fails to react with sialic acid-specific stains (Jackson, 1978). The proportion of carbohydrate in the particle is not known.
Lipid: No information, but density of the particles and analogy to other enveloped viruses suggest that the lipid composition is probably 15-20%
Enzyme activity: Purified preparations contain a RNA-dependent RNA polymerase as is also found for lettuce necrotic yellows, broccoli necrotic yellows and sonchus viruses (Peters et al., 1978). A similar enzyme present in a nucleocapsid preparation from infected plants produced a RNA transcript that cross-hybridized with about 40% of the viral RNA sequence (D. Peters, personal communication).
Relations with Cells and Tissues
Nuclear inclusions (Fig. 7) are found in leaf cells infected with sonchus yellow net virus in epidermal strips examined in the light microscope (Christie et al., 1974; Christie & Edwardson, 1978). Many particles are present in the nucleus or perinuclear space and a few particles are found in the cytoplasm of plant cells (Fig. 8). Bacilliform particles are also found in cells of viruliferous aphids (Christie et al., 1974). RNA molecules complementary to the RNA in the virus particles are found in infected tobacco cells (Milner & Jackson, 1979). These are presumed to be messenger RNA transcripts because they are bound to polyribosomes, are composed of polyadenylated and non-polyadenylated species and are similar in size to transcripts of animal rhabdoviruses (Wagner, 1975). These complementary RNA sequences are associated with both free and membrane-bound polyribosomes (J. J. Milner, M. J. J. Hakkaart & A. O. Jackson, unpublished results).
Notes
Five plant rhabdoviruses are known to infect lettuce and sowthistle. Of these, sonchus yellow net virus differs from lettuce necrotic yellows virus (Francki & Randles, 1970) in host range, aphid vectors, serology, particle size and protein composition; from sowthistle yellow vein virus (Peters, 1971) in host range, aphid vectors, serology and mechanical transmissibility; from sonchus virus (Vega et al., 1976) in host range and protein composition; and from gomphrena virus (Kitajima & Costa, 1966) in host range. Another virus infecting sowthistle, sonchus yellow vein virus, is reportedly non-persistent in aphids, and thus not likely to be a rhabdovirus (Reddy & Janardhan, 1975).
Figures

Electron micrograph of a leaf dip preparation from infected Nicotiana hybrid, negatively stained with potassium phosphotungstate. Bar represents 200 nm.

Local lesions on mechanically inoculated Chenopodium quinoa (12 days after inoculation).

Systemic symptoms on mechanically inoculated Nicotiana hybrid (14 days after inoculation).

Symptoms on mechanically inoculated sowthistle (Sonchus oleraceus). (Left to right): systemic net pattern, chlorotic local lesions, healthy leaf.

Electron micrograph of particles from a purified virus preparation fixed with gluraraldehyde and stained with potassium phosphotungstate. Bar represents 100 nm.

Polyacrylamide gel electrophoresis of proteins from sodium dodecyl sulphate-disrupted particles of sonchus yellow net virus (SYNV) and potato yellow dwarf virus (PYDV). G = glycoprotein, N = nucleic acid-associated protein, M1 and M2 = membrane matrix proteins.
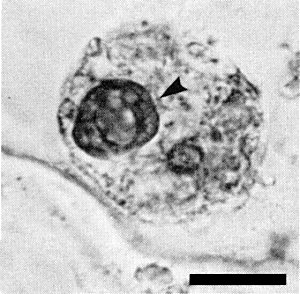
Viral inclusion (arrow) in the nucleus of an infected epidermal cell of Nicotiana hybrid stained with Azure A. The smaller staining body is the nucleolus. Bar represents 10 µm. (Photograph kindly supplied by Mr. R. G. Christie.)

Thin section of an infected tobacco cell with clumps of virus particles in the nucleus near its periphery. Bar represents 1 µm.
References list for DPV: Sonchus yellow net virus (205)
- Black, CMI/AAB Descriptions of Plant Viruses 35, 4 pp., 1970.
- Christie, Pl. Dis. Reptr 53: 939, 1969.
- Christie & Edwardson, A. Rev. Phytopath. 16: 31, 1978.
- Christie, Christie & Edwardson, Phytopathology 64: 840, 1974.
- Fenner, Intervirology 7: 1, 1976.
- Francki & Randles, CMI/AAB Descriptions of Plant Viruses 26, 4 pp., 1970.
- Jackson, Virology 87: 172, 1978.
- Jackson & Christie, Virology 77: 344, 1977.
- Kitajima & Costa, Virology 29: 523, 1966.
- Milner & Jackson, Virology 97: 90, 1979.
- Peters, CMI/AAB Descriptions of Plant Viruses 62, 3 pp., 1971.
- Peters, Toriyama & Terlouw, Abstr. 4th int. Congr. Virol. 405, 1978.
- Reddy & Janardhan, Curr. Sci. 44: 596, 1975.
- Vega, Gracia, Rubio-Huertos & Feldman, Phytopath Z. 85: 7, 1976.
- Wagner, Comprehensive Virology 4: 1, 1975.