Details of DPV and References
DPV NO: 212 July 1979
Family: Tombusviridae
Genus: Carmovirus
Species: Cowpea mottle virus | Acronym: CPMoV
Cowpea mottle virus
R. F. Bozarth Department of Life Sciences, Indiana State University, Terre Haute, Indiana 47809, USA
S. A. Shoyinka University of Ife, Moor Plantation, P.M.B. 5029, Ibadan, Nigeria
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
Described by Robertson (1966).
A virus with RNA-containing isometric particles about 30 nm in diameter which sediment as a single component. It has a wide host range, is transmitted mechanically and by beetles, and is known only from Nigeria.
Main Diseases
Causes mottling or bright yellow mosaic in cowpea (Vigna unguiculata) (Fig. 1). Leaves are distorted, reduced in size and a witches’ broom syndrome is common. Yields may be reduced by more than 75%. Field-grown plants are often also infected with cowpea mosaic or southern bean mosaic viruses. In naturally infected Bambarra groundnut (Voandzeia subterranea) disease symptoms include green mottle, stunting, and shortening of petioles.
Geographical Distribution
Reported only from Nigeria.
Host Range and Symptomatology
Species of only a few plant families have been tested. The virus gives a systemic mosaic in many members of the Leguminosae. Several members of the Solanaceae become systemically infected without showing symptoms.
-
Diagnostic species
- Chenopodium amaranticolor
and C. quinoa. Diffuse chlorotic local lesions (Fig. 3). No systemic infection. - Vigna unguiculata (cowpea). Symptoms vary with environment and with the variety tested.
Initial symptoms may be diffuse chlorotic or necrotic spots on inoculated leaves, followed by intense
chlorosis of the entire lamina or extensive veinal necrosis. When young primary leaves are inoculated,
the first trifoliolate leaf is severely affected, with severe veinal necrosis at the base. Inoculation
of primary leaves later (2 weeks after sowing) results in a less severe reaction. In either case,
expanding trifoliolate leaves develop a severe chlorotic mottle
(Fig. 1) or green mottle (Fig. 2),
and are distorted and reduced in size.
- Cajanus cajan (pigeon pea). Systemic symptoms similar to those in cowpea (Fig. 3).
- Phaseolus vulgaris, P. mungo, P. lunatus, P. acutifolius, P. aureus, P. lathyroides and Glycine max develop systemic mosaic.
- Cajanus cajan (pigeon pea). Systemic symptoms similar to those in cowpea (Fig. 3).
-
Propagation species
- Vigna unguiculata
is suitable for propagation and maintenance of the virus. The cv. California No. 5 (USA) or Ife Brown (Nigeria) have been used, but others are probably just as good.Assay species
- Chenopodium amaranticolor
and C. quinoa are useful local lesion hosts. Vigna unguiculata is satisfactory under some conditions. Robertson (1966) reported that Phaseolus vulgaris cv. Masterpiece develops necrotic local lesions.
Strains
Isolates from Voandzeia subterranea and Vigna unguiculata differ somewhat in host range but not serologically (Shoyinka et al., 1978).
Transmission by Vectors
Transmitted principally by the galerucid beetle, Ootheca mutabilis (Shoyinka et al., 1978) and also by Paraluperodes lineata (D. J. Allen, unpublished data). O. mutabilis acquired virus in 10 min, transmitted within 1 h, and remained infective for 5 days. The virus was recovered from faeces of adult beetles that had fed on infected plants (S. A. Shoyinka, unpublished data).
Transmission through Seed
The virus may be seed-borne in cowpea (cv. Ibadan White, California No. 5), Bambarra groundnut and French bean. The virus is transmitted to seedlings through up to 10% of the seeds of diseased cowpea plants, depending on the cultivar and the interval of time between infection and flowering (Shoyinka et al., 1978).
Serology
The virus is a good immunogen. Antiserum with a titre of 1/640 was obtained from a rabbit given 2 injections of 2 mg virus. Antiserum diluted 1/5 in 0.85% NaCl and 50% glycerol was still reactive after 2 years’ storage at 25-28°C. A single line of precipitate is obtained in agar double-diffusion tests.
Relationships
Cowpea mottle virus has a combination of properties unlike those of any previously described virus. Purified virus did not react with antiserum to 19 isometric viruses; none of 16 isometric viruses reacted with cowpea mottle virus antiserum (Shoyinka et al., 1978).
Stability in Sap
In crude sap of cowpea, the virus was infective after dilution more than 2 x 10-6, or after heating for 10 min at 60°C but not 65°C, but lost infectivity after incubation for 24 h at 22°C.
Purification
Systemically infected leaves of cowpea cv. California No. 5 were harvested 2 to 3 weeks after inoculation of primary leaves, blended in 0.1 M potassium phosphate buffer, pH 7.0, and subjected to three cycles of differential centrifugation. All sediments were resuspended in this buffer. Finally, purification was by sucrose density gradient centrifugation (Shoyinka et al., 1978).
Properties of Particles
(Shoyinka et al., 1978) Purified preparations contain a single sedimenting component.
Sedimentation coefficient: s°20,w = 122 S.
Mol. wt: estimated from percentage RNA to be about 7.0 x 106.
Absorbance at 260 nm (1 mg/ml, 1 cm light path): about 5.
A260/A280: 1.66.
Buoyant density in CsCl: 1.349 ± 0.004.
Particle Structure
Isometric, 30 ± 1.5 nm when stained with 1% uranyl acetate (Fig. 4).
Particle Composition
Nucleic acid: RNA, single-stranded. M. Wt: 1.4 x 106 estimated by electrophoresis in 2.4% polyacrylamide gels using RNA from cowpea mosaic virus and tobacco mosaic virus as markers (D. K. Edge & R. F. Bozarth, unpublished data). Nucleic acid is approximately 20% of particle weight (estimated from buoyant density in CsCl).
Protein: Subunit M. Wt = 44,500 (estimated from electrophoresis in polyacrylamide gels) (Shoyinka et al., 1978).
Relations with Cells and Tissues
Virus has been recovered from leaves, whole flowers, anthers, pistils, whole immature pods, and milk stage seed, but not from the flower keel of systemically infected plants.
Notes
This virus has not been reported outside Nigeria although its occurrence in other tropical countries is suspected. It has not been detected in limited surveys in Puerto Rico, El Salvador, Colombia and Costa Rica, and there is no evidence that it occurs in the USA. Every effort should be made to prevent its spread throughout the world, especially in contaminated seed.
The virus is extremely virulent in cowpea and has been known to spread to uninoculated plants in the absence of a known vector. Powdery mildew is suspected to play a role in such spread, but no definitive experiments have been done.
The identification of cowpea mottle virus without the use of serology is difficult. In its general symptomatology it resembles other cowpea viruses, especially cowpea mosaic (Van Kammen & De Jager, 1978), cowpea severe mosaic (De Jager, 1979) and cowpea chlorotic mottle (Bancroft, 1971) viruses. Multiple infections with cowpea mosaic virus and southern bean mosaic virus are common throughout its known geographical distribution. Cowpea mottle virus tends to produce a brilliant chlorotic mottle in most susceptible legumes, many of which are non-hosts or symptomless hosts of cowpea mosaic and cowpea chlorotic mottle viruses. Also cowpea mottle virus differs from cowpea mosaic virus in not infecting Chenopodium amaranticolor systemically.
Figures

Systemically infected leaves of cowpea cv. Brabham 892A showing chlorosis and mottle symptoms (left) or severe chlorosis (centre). A comparable leaf from a non-inoculated plant is shown on the right.

Green mottle symptoms in trifoliolate leaf of Vigna unguiculata cv. Ife Brown.

(Above) Diffuse chlorotic spots in infected leaf (left) and healthy leaf (right) of Chenopodium amaranticolor. (Below) Brilliant yellow blotch symptoms in trifoliolate leaf (left) and healthy leaf (right) of Cajanus cajan.
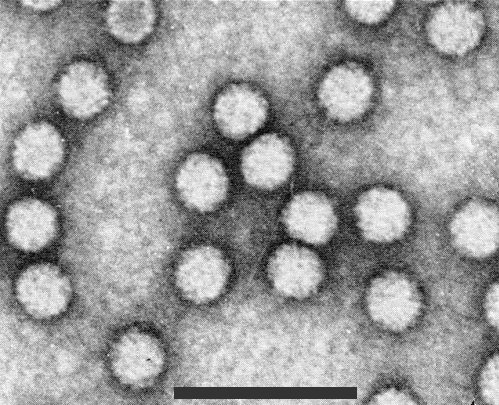
Electron micrograph of particles negatively stained with 1% uranyl acetate. Bar represents 100 nm.
References list for DPV: Cowpea mottle virus (212)
- Bancroft, CMI/AAB Descriptions of Plant Viruses 49, 4 pp., 1971.
- De Jager, CMI/AAB Descriptions of Plant Viruses 209, 5 pp., 1979.
- Robertson, B.Sc. Thesis, Univ. of Oxford, England, 1966.
- Shoyinka, Bozarth, Reese & Rossel, Phytopathology 68: 693, 1978.
- Van Kammen & De Jager, CMI/AAB Descriptions of Plant Viruses 197, 6 pp., 1978.