Details of DPV and References
DPV NO: 214 July 1979
Family:
Genus:
Species: | Acronym:
Tymovirus group
Renate Koenig Institut für Viruskrankheiten der Pflanzen Braunschweig, Germany
D.-E. Lesemann Institut für Viruskrankheiten der Pflanzen Braunschweig, Germany
Contents
- Type Member
- Main Characteristics
- Members
- Geographical Distribution
- Transmission by Vectors
- Ecology and Control
- Relations with Cells and Tissues
- Properties of Particles
- Genome Properties
- Replication
- Satellite
- Defective-Interfering RNA
- Relationships
- Notes on Tentative Members
- Affinities with Other Groups
- Notes
- Acknowledgements
- Figures
- References
Type Member
Turnip yellow mosaic virus.
Main Characteristics
Two major classes of isometric particle, c. 30 nm in diameter with hexagonal outlines, sedimenting at c. 115 and 54 S and containing respectively c. 35 and 0-2.5% single-stranded RNA. Genomic RNA, M. Wt c. 2 x 106, in bottom component (‘full particles’); sub-genomic coat protein messenger RNA, M. Wt 200,000-300,000, either in top (‘shells’) or bottom component. Each particle contains 180 molecules of a single coat polypeptide, M. Wt c. 20,000. Thermal inactivation point 65-95°C; longevity in sap a few weeks; concentration in sap often 50-500 mg/l. Host range usually narrow. Symptoms are mottle or bright yellow mosaic. Virus particles are found in cytoplasm (‘full particles’ and ‘shells’), in nuclei (‘shells’), and rarely in chloroplasts and mitochondria. Chloroplasts develop small peripheral vesicles bounded by double membranes, and become rounded and clumped. Transmissible by inoculation with sap; beetle vectors are known for several members.
Recent reviews on tymoviruses are by Matthews (1977) and Koenig & Lesemann (1981).
Members
Table 1 lists the definitive members of the group together with some of their properties.
Table 1. Definitive members of the tymovirus group
| Virus | Description no. or ref |
Geographical distribution |
Families with species susceptible to systemic infection |
Reported beetle vector |
| Andean potato latent (APLV) | 124, (a) | South American Andes | Solanaceae, Chenopodiaceae | Epitrix sp. |
| Belladonna mottle (BMV) | 52 | Europe | Solanaceae | Epitrix atropae |
| Cacao yellow mosaic (CoYMV) | 11 | Africa (Sierra Leone) | Sterculiaceae, Apocynaceae, Bombacaceae, Chenopodiaceae, Solanaceae | None reported |
| Clitoria yellow vein (CYVV) | 171 | Africa (Kenya) | Leguminosae, Malvaceae, Solanaceae | None reported |
| Desmodium yellow mottle (DeYMV) | 168 | USA | Leguminosae | None reported |
| Dulcamara mottle (DMV) | (b) | Europe | Solanaceae | Psylliodes affinis |
| Eggplant mosaic (EMV) | 124 | Trinidad, strains in North and South America | Solanaceae, Chenopodiaceae | Epitrix sp. |
| Erysimum latent (ELV) | 222, (c) | Europe (Germany) | Amaranthaceae, Caryophyllaceae, Cruciferae, Labiatae, Leguminosae and Resedaceae | Phyllotreta spp. |
| Kennedya yellow mosaic (KYMV) | 193 | Australia | Leguminosae, Solanaceae | None reported. |
| Okra mosaic (OkMV) | 128 | Africa (Ivory Coast, Nigeria) | 23 families including all those listed for the other viruses except Valerianaceae and Caesalpiniaceae | Podagrica decolorata |
| Ononis yellow mosaic (OYMV) | (b) | Europe (England) | Leguminosae, Solanaceae | None reported |
| Physalis mosaic (PhyMV) | (d) | USA | Solanaceae | None reported |
| Plantago mottle (PlMV) | (e) | USA | Plantaginaceae, Aizoaceae, Leguminosae, Scrophulariaceae, Solanaceae | None reported |
| Scrophularia mottle (ScrMV) | 113 | Europe (Germany) | Scrophulariaceae, Caryophyllaceae, Labiatae, Solanaceae, Umbelliferae and Valerianaceae | Cionis spp. |
| Turnip yellow mosaic (TYMV) | 2, 230 | Europe | Cruciferae, Resedaceae | Phyllotreta spp., Psylliodes spp. |
| Wild cucumber mosaic (WCuMV) | 105 | USA | Cucurbitaceae | Acalymma trivittata |
(a) Koenig, Fribourg & Jones, 1979; (b) Gibbs et al., 1966; (c) Shukla, Proeseler & Schmelzer, 1975; (d) Peters & Derks, 1974; (e) Granett, 1973.
Geographical Distribution
Tymoviruses have been reported from most parts of the world including tropical, subtropical and temperate regions. Restricted host ranges (Table 1), frequently in wild plants, and lack of vector insects are probably the main reasons for the limited distribution of individual tymoviruses.
Transmission by Vectors
Several tymoviruses are transmitted by beetles in the families Chrysomelidae and Curculionidae. The efficiency of transmission is low with some tymoviruses (e.g. ScrMV, APLV) and high with others (TYMV, BMV). The former viruses may be transmitted by a purely mechanical process (Desc. 113; Jones & Fribourg, 1977) whereas the latter may become more intimately associated with the beetles (Walters, 1969; Weidemann & Bode, 1973). The viruses are retained by the insects for a few hours (ScrMV) up to several days (BMV, TYMV).
A single instance of whitefly transmission of a tymovirus (OkMV) has been reported (Lana & Taylor, 1976). No other vectors of biological importance are known.
Ecology and Control
Tymoviruses are highly infectious and are spread by mechanical contact, agricultural machinery, beetle vectors and probably on the bodies of contaminated animals. True seed transmission has been recorded for APLV (Desc. 124). Most natural host plants are perennials, and this aids the survival of tymoviruses.
Relations with Cells and Tissues
Tymoviruses invade all main tissues of their host plants. Effects on cells have been studied almost exclusively in leaf parenchyma tissues. Characteristic changes are induced in the fine structure of chloroplasts (for reviews see Matthews, 1973; Lesemann, 1977), notably the formation of small vesicles at the periphery, bounded by double membranes (50-100 nm thick) which seem to be continuous with the chloroplast double membrane (Hatta & Matthews, 1974; Hatta, Bullivant & Matthews, 1973). The vesicles have a narrow neck and often contain fibrils thought to represent double-stranded nucleic acid. The chloroplasts become clumped and virus particles accumulate in the cytoplasm. Later in infection the chloroplasts develop large internal vesicles or ‘vacuoles’ which may at least in part originate from inflated small peripheral vesicles (Lesemann, 1977). The ‘vacuoles’ may be seen with the light microscope in TYMV-infected protoplasts; their formation depends on an intact photosynthetic apparatus and on exposure to light (Matthews & Sarkar, 1976). After the bulk of virus has been produced, further changes may lead to a more or less complete disorganization of the chloroplasts.
Virus particles are first found in electron-lucent zones which form in the cytoplasm over the chloroplast vesicles. Virus particles eventually reach high concentrations in the cytoplasm, and the cell vacuoles are also often invaded as a result of breakages of the tonoplast. Very seldom, particles are found within chloroplasts and mitochondria (Lesemann, 1977). Under suitable conditions, e.g. wilting or plasmolysis (Hatta & Matthews, 1974; Hatta, 1976) virus particles form crystalline aggregates in the cytoplasm, nuclei and vacuoles (Lesemann, 1977). With all tymoviruses studied, ‘shells’ accumulate in the nuclei early in infection and may later fill a major part of the nuclear space. They are, however, also present in the cytoplasm (Hatta & Matthews, 1976; Lesemann, 1977).
In addition to the described group-specific cytopathic effects, virus- or strain-specific changes may be induced which are discussed in detail by Lesemann (1977) and Koenig & Lesemann (1981).
Properties of Particles
Tymoviruses have a single species of coat protein of c. 20,000 M. Wt. The amino acid composition of tymoviruses (H. L. Paul, A. J. Gibbs & B. Wittmann-Liebold, unpublished results) and the complete amino acid sequence of TYMV (Peter et al., 1972) are known. The protein shells are T = 3 icosahedral structures containing 180 protein subunits clustered into 12 pentamers and 20 hexamers (Klug, Longley & Leberman, 1966). In sucrose density gradients, two major and in some instances several minor components are separated. The biological significance of the minor components is uncertain (Matthews, 1977). Particles of the major ‘bottom’ component sedimenting at c. 115 S contain one molecule of single-stranded RNA of M. Wt 2 x 106 which accounts for about 35% of the particle weight. In TYMV the ‘bottom’ component (Szybiak, Bouley & Fritsch, 1978) also contains a sub-genomic RNA of M. Wt c. 0.25 x 106 which acts as coat protein messenger (Klein et al., 1976; Pleij et al., 1976). The major ‘top’ component sedimenting at c. 54 S contains no RNA in TYMV and coat protein mRNA in EMV, OkMV and WCuMV (Szybiak et al., 1978). The ‘top’ component of EMV also contains transfer RNA species of plant origin (Pinck & Hall, 1978). Spermidine and spermine account for 1% and less than 0.04%, respectively, of the particle weight of TYMV (Beer & Kosuge, 1970) and may be important in neutralizing some of the phosphate groups of the RNA.
Particles of tymoviruses are stabilized by hydrophobic protein-protein interactions (Kaper, 1975) which are stronger in TYMV than in EMV (Jonard et al., 1976; Bouley, Briand & Witz, 1977). The presence of RNA destabilizes the protein-protein interactions (Jonard et al., 1976; Bouley et al., 1977) and leads to conformational changes in the protein (Tamburro et al., 1978). Protein-RNA interactions probably involve hydrogen bonds between protein carboxylate and nucleotide amino groups (Kaper, 1975; Jonard et al., 1976). In attempts to reassemble nucleic acid-containing particles of TYMV in vitro, virus-like protein-RNA complexes were obtained at low pH in the presence of magnesium or spermidine but these were unstable at neutral pH (Jonard et al., 1976). Similar complexes were also obtained with TYMV protein and RNA species from EMV or BMV, but not with RNA from tobacco mosaic virus (Bouley et al., 1975).
Genome Properties
The reported base compositions of tymovirus RNA species are within the range of G 15.2-17.5 : A 17.0-23.8 : C 31.9-42.1 : U 22.1-29.4. Cytidine is always the most common nucleotide.
The nucleotide sequence of the genomic RNA of M. Wt 2 x 106 encodes all the information for viral infectivity, and includes the cistron for the coat protein, which is located at its 3' extremity. In cell-free systems the genomic RNA directs the synthesis of one or more large proteins, but not of viral coat protein. This is done by the sub-genomic viral coat protein messenger RNA of M. Wt c. 0.25 x 106 which is derived from the genomic RNA in vivo in an unknown way (Klein et al., 1976; Pleij et al., 1976; Ricard et al., 1977; Szybiak et al., 1978). The entire nucleotide sequence is known for the coat protein messenger RNA of TYMV (Guilley & Briand, 1978); the sequence of 567 nucleotides in the coat protein cistron predicts the amino acid sequence actually found by Peter et al. (1972). The genomic and sub-genomic RNA molecules both have 5' terminal 7-methylguanosine (Briand, Keith & Guilley, 1978) and both have at their 3' ends a non-translated sequence of 109 nucleotides which can act as a substrate for various tRNA-specific enzymes, including valyl-tRNA synthetase (see Briand et al., 1977, for review). Both RNA molecules have the same base composition (Guilley & Briand, 1978).
Replication
The small vesicles induced by tymoviruses in the periphery of chloroplasts contain membrane-bound viral replicase (Laflèche et al., 1972) and are probably the main site of production of viral plus-strand RNA. Various aspects of TYMV replication have been discussed in detail by Hatta & Matthews (1976).
Relationships
Tymoviruses have been classified according to RNA base composition (Gibbs et al., 1966), serological interrelationships (Fig. 1), cytopathic effects (Lesemann, 1977) and coat protein amino acid composition (H. L. Paul, A. J. Gibbs & B. Wittmann-Liebold, unpublished results). Serologically, several tymoviruses are closely related and some might be regarded as strains of the same virus, e.g. OYMV and PlMV as strains of ScrMV, APLV as a strain of EMV, and DMV as a strain of BMV (Desc. 124; Koenig, 1976). Perhaps all the tymoviruses isolated from solanaceous hosts, i.e. the many different strains of APLV, EMV, PhyMV, BMV and DMV, might be considered as strains of one virus. The divergent views on this question have been discussed by Koenig & Lesemann (1981).
Affinities with Other Groups
No close affinities of tymoviruses with viruses in other groups are known. It is not known whether or not the weak serological cross reactivities of several tymoviruses with cocksfoot mild mosaic and carnation ringspot viruses reflect true relationships or chance similarities (Bercks & Querfurth, 1972).
Acknowledgements
The authors’ work on tymoviruses was supported by the Deutsche Forschungsgemeinschaft.
Figures
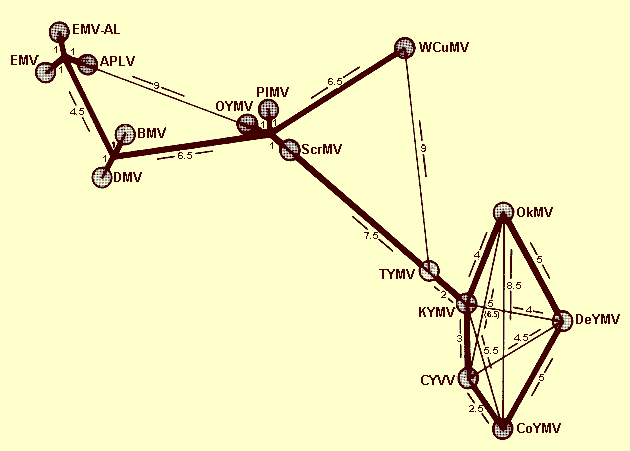
Serological classification of tymoviruses on the basis of average serological differentiation indices (SDI) in reciprocal tests. The SDI is the number of two-fold dilution steps separating heterologous and homologous titres of an antiserum. The average SDI of reciprocal tests are depicted as length units. The scheme is based on more than 4000 individual titre determinations with 321 bleedings from 56 rabbits. For details see Koenig (1976).
References list for DPV: Tymovirus group (214)
- Beer & Kosuge, Virology 40: 930, 1970.
- Bercks & Querfurth, Phytopath. Z. 75: 215, 1972.
- Bouley, Briand, Jonard, Witz & Hirth, Virology 63: 312, 1975.
- Bouley, Briand & Witz, Virology 78: 425, 1977.
- Briand, Jonard, Guilley, Richards & Hirth, Eur. J. Biochem. 72: 453, 1977.
- Briand, Keith & Guilley, Proc. natn. Acad. Sci. U.S.A. 75: 3168, 1978.
- Gibbs, Hecht-Poinar, Woods & McKee, J. gen. Microbiol. 44: 177, 1966.
- Granett, Phytopathology 63: 1313, 1973.
- Guilley & Briand, Cell 15: 113, 1978.
- Hatta, Virology 69: 237, 1976.
- Hatta & Matthews, Virology 59: 383, 1974.
- Hatta & Matthews, Virology 73: 1, 1976.
- Hatta, Bullivant & Matthews, J. gen Virol. 20: 37, 1973.
- Jonard, Briand, Bouley, Witz & Hirth, Phil. Trans. R. Soc. Ser. B 276: 123, 1976.
- Jones & Fribourg, Ann. appl. Biol. 86: 123, 1977.
- Kaper, The Chemical Basis of Virus Structure, Dissociation and Reassembly, Amsterdam: North Holland, 485 pp., 1975.
- Klein, Fritsch, Briand, Richards, Jonard & Hirth, Nucleic Acids Res. 3: 3043, 1976.
- Klug, Longley & Leberman, J. molec. Biol. 15: 315, 1966.
- Koenig, Virology 72: 1, 1976.
- Koenig & Lesemann, in Handbook of Plant Virus Infections and Comparative Diagnosis, p. 33, ed. Kurstak, Amsterdam: Elsevier/North-Holland, 1981.
- Koenig, Fribourg & Jones, Phytopathology, 69: 748, 1979.
- Laflèche, Bové, Dupont, Mouchés, Astier, Gamier & Bové, in Proc. 8th Meet. Fed. Eur. biochem. Soc., Amsterdam 1972, pp. 43-71, Amsterdam: North-Holland, 1972.
- Lana & Taylor, Ann. appl. Biol. 82: 361, 1976.
- Lesemann, Phytopath. Z. 90: 315, 1977.
- Matthews, A. Rev. Phytopath. 11: 147, 1973.
- Matthews, in The Atlas of Insect and Plant Viruses, p. 347, ed. K. Maramorosch, New York: Academic Press, 1977.
- Matthews & Sarkar, J. gen. Virol. 33: 435, 1976.
- Peter, Stehelin, Reinbolt, Collot & Duranton, Virology 49: 615, 1972.
- Peters & Derks, Neth. J. Pl. Path. 80: 124, 1974.
- Pinck & Hall, Virology 88: 281, 1978.
- Pleij, Neeleman, Van Vloten-Doting & Bosch, Proc. natn. Acad. Sci. U.S.A. 73: 4437, 1976.
- Ricard, Barreau, Renaudin, Mouchés & Bové Virology 79: 231, 1977.
- Shukla, Proeseler & Schmelzer, Acta phytopath. Acad. Sci. hung. 10: 211, 1975.
- Szybiak, Bouley & Fritsch, Nucleic Acids Res. 5: 1821, 1978.
- Tamburro, Guantieri, Piazzolla & Gallitelli, J. gen. Virol. 40: 337, 1978.
- Walters, Adv. Virus Res. 15: 339, 1969.
- Weidemann & Bode, Phytopath. Z. 76: 6, 1973.