Details of DPV and References
DPV NO: 218 September 1980
Family: Potyviridae
Genus: Potyvirus
Species: Carrot thin leaf virus | Acronym: CTLV
Carrot thin leaf virus
W. E. Howell Washington State University, Irrigated Agriculture Research and Extension Center, Prosser, WA 99350, USA
G. I. Mink Washington State University, Irrigated Agriculture Research and Extension Center, Prosser, WA 99350, USA
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
Described by Howell & Mink (1976).
A virus with filamentous particles c. 736 x 11 nm, infecting several species of Umbelliferae and a few species from other families. It is transmitted by aphids in a non-persistent manner and is sap-transmissible.
Main Diseases
Causes twisting and narrowing of leaves of carrot and induces yield losses of nearly 25%.
Geographical Distribution
Reported only from the semi-desert irrigated areas of north-western USA.
Host Range and Symptomatology
Species from three dicotyledonous families (Umbelliferae, Chenopodiaceae and Solanaceae) are susceptible to infection. Some species of Umbelliferae become infected only by aphid inoculation, but other hosts are readily infected by inoculation of sap.
- Diagnostic species
- Coriandrum sativum
(coriander) and Daucus carota ssp. sativa (carrot). Leaflet lobes of newly formed leaves become twisted and narrow 2-3 weeks after inoculation. Vein clearing, chlorotic spots and faint mottling also occur (Fig. 1). Carrot is readily infected only by aphid inoculation. - Nicotiana clevelandii. Faint chlorotic mottle in young uninoculated leaves.
- Chenopodium quinoa. Chlorotic local lesions 5-7 days after inoculation. The lesions may later develop necrotic centres (Fig. 2).
- Chenopodium quinoa. Chlorotic local lesions 5-7 days after inoculation. The lesions may later develop necrotic centres (Fig. 2).
- Propagation species
- Carrot is suitable for maintaining cultures, C. quinoa is a good source of virus for
purification.
- Assay species
- C. quinoa
and C. amaranticolor are good local lesion assay hosts. Coriander and carrot are suitable for assaying transmission by vectors.
Strains
None recorded.
Transmission by Vectors
Transmitted in the non-persistent manner by the aphids Myzus persicae and Cavariella aegopodii. Aphids can acquire the virus in 5-10 min feeding periods. The virus is spread rapidly in the field, apparently by M. persicae (Howell & Mink, 1977).
Transmission through Seed
No transmission through carrot seed has been detected (Howell & Mink, 1979).
Serology
The virus appears to be only weakly immunogenic. A rabbit given four intravenous injections each of 0.5 mg virus, gave an antiserum with a homologous titre of only 1/64 in ring precipitin tests. The virus can be detected in sap of infected plants by electron microscope serology (Milne & Luisoni, 1977).
Relationships
The virus has many properties in common with members of the potyvirus group, but in ring-precipitin tests it was serologically unrelated to three potyviruses that infect umbelliferous plants (clover yellow vein virus, parsnip mosaic virus, and the type, poison hemlock and parsley strains of celery mosaic virus) or to ten other potyviruses (bean common mosaic, bean yellow mosaic, iris mild mosaic, peanut mottle, pea seed-borne mosaic, potato virus Y, soybean mosaic, sugarcane mosaic, tobacco etch and turnip mosaic viruses).
Stability in Sap
In C. quinoa sap the thermal inactivation point (10 min) is 50-55°C, dilution end point is beyond 10-5 and longevity in vitro at 22°C is 2 days.
Purification
The virus is stable and is readily purified from senescent inoculated leaves of C. quinoa by the following method. Blend each 50 g of frozen, infected leaves in 150 ml of neutral 0.01 M potassium phosphate buffer. Squeeze the extract through cheesecloth and emulsify with an equal volume of chloroform by stirring for 30 min. Break the emulsion by centrifugation at 3000 g for 30 min, and centrifuge the aqueous phase for 1.5 h at 71,000 g. Resuspend the sediment in 0.01 M phosphate buffer, pH 7.0. After low speed centrifugation, centrifuge the virus through a 10 ml cushion of 30% sucrose containing 4% polyethylene glycol (M. Wt 6000) and 0.12 M NaCl at 60,000 g for 2 h in a swinging bucket rotor. Further purification is obtained by centrifugation in sucrose density gradients. Yield of virus is about 20 mg/kg leaf.
Properties of Particles
A260/A280: 1.18.
Particle Structure
Particles are flexuous filaments c. 736 x 11 nm (Fig. 3).
Particle Composition
Nucleic acid content is c. 4% (based on A260/A280 ratio).
Relations with Cells and Tissues
No information.
Notes
Weed hosts (wild Daucus carota and Conium maculatum), regrowth from discarded carrot plants, overlapping carrot cropping seasons, and infestations of green peach aphids, all contribute to epidemics of carrot thin leaf disease in Washington State (Howell & Mink, 1977). In the main carrot growing areas of central Washington nearly all the carrots in some fields contain the virus by harvest time. Carrot thin leaf virus has similarities in morphology and host range to celery mosaic, parsnip mosaic, carrot mosaic and clover yellow vein viruses but is distinguished from them by its symptoms on carrot and Nicotiana clevelandii.
Figures

Leaves from (left) infected and (right) healthy Daucus carota ssp. sativa.

Chlorotic local lesions in inoculated leaves of Chenopodium quinoa.
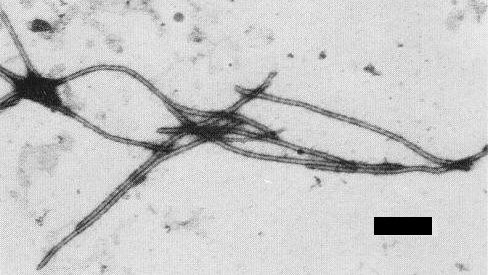
Virus particles in uranyl acetate. Bar represents 185 nm.
References list for DPV: Carrot thin leaf virus (218)
- Howell & Mink, Phytopathology 66: 949, 1976.
- Howell & Mink, Pl. Dis. Reptr 61: 841, 1977.
- Howell & Mink, Pl. Dis. Reptr 63: 989, 1979.
- Milne & Luisoni, in Methods in Virology, Vol. 6, p. 265, ed. K. Maramorosch & H. Koprowski, London and New York: Academic Press, 1977.