Details of DPV and References
DPV NO: 219 September 1980
Family: Caulimoviridae
Genus: Caulimovirus
Species: Strawberry vein banding virus | Acronym: SVBV
Strawberry vein banding virus
N. W. Frazier Division of Nematology, University of California, Davis, California 95616, USA
R. H. Converse USDA, SEA-AR, Department of Botany & Plant Pathology, Oregon State University, Corvallis, Oregon 97331, USA
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
Described by Frazier (1955).
- Selected synonyms
- Yellow vein banding virus (Rev. appl. Mycol. 38: 217)
- Chiloensis vein banding virus (Rev. appl. Mycol. 39: 725)
- Eastern vein banding virus (Rev. appl. Mycol. 39: 725)
- Chiloensis vein banding virus (Rev. appl. Mycol. 39: 725)
A virus with isometric particles 40 to 50 nm in diameter. The particles frequently occur in cytoplasmic inclusions. The virus is transmissible in a semi-persistent manner by several species of aphid, by dodder, and by grafting, but not by inoculation of sap. It appears to be of minor economic importance because of low incidence, but can be very damaging when in mixed infections with other viruses, such as strawberry crinkle.
Main Diseases
Causes vein banding in strawberry (Fig. 1); in complexes with other viruses, it may induce variations of crinkle, yellows and leaf curl diseases (Plakidas, 1964; Miller & Frazier, 1970).
Geographical Distribution
Native to temperate regions of eastern and western North America; occurs in Australia and Brazil, probably as the result of introduction in planting material, and possibly in Europe (Schöniger, 1958).
Host Range and Symptomatology
Has a narrow natural host range in the genus Fragaria, principally in the wild species, F. chiloensis, F. vesca and F. virginiana, and in the cultivated species, F. ananassa. Sanguisorba minor has been experimentally infected by aphid transmission and by leaf grafting (Morris et al., 1980). The virus is transmitted readily by aphid vectors (Frazier, 1955; Mellor & Forbes, 1960), grafting (Frazier, 1974a) and dodder (Frazier, 1955), but not by inoculation with sap.
-
Diagnostic species
- Fragaria vesca
clone UC-6 (Frazier, 1974b) and F. virginiana clone UC-12 (Frazier, 1974b): mild leaf curl (epinasty of midribs and petioles, and twisting of leaflets) are the first symptoms but the most diagnostic is discontinuous chlorotic banding of the main veins (Fig. 1). Later there is necrosis and stunting, which may be severe. In mixed infection with strawberry crinkle virus, there is severe and chronic leaf curl.Propagation species
- Cultures may be propagated satisfactorily in most species and clones of Fragaria (e.g. F. ananassa, F. chiloensis, F. vesca and F. virginiana).
-
Assay species
- Fragaria vesca
clone UC-6 and F. virginiana clone UC-12 are the currently preferred test clones. F. vesca var. semperflorens (the seed-propagated Alpine strawberry) shows symptoms when infected with most variants of the virus and is very useful for virus assay by vector transmission (Frazier, 1974b).
Strains
Strains have been distinguished by severity of symptoms and by aphid transmissibility (Frazier & Posnette, 1958; Frazier, 1960). Their distribution and importance are not known. The type strain (Frazier, 1955) has been used experimentally more than others. Erdbeernekrose-Virus (Schöniger, 1958) may be a strain, but this requires confirmation. Incubation periods required for symptom development in indicator plants are 3-5 weeks for the leaf-curl strain (Prentice, 1952), 2-3 weeks for the vein banding strain (Frazier, 1955), and 3-4 weeks for Erdbeernekrose-Virus (Schöniger, 1958).
Transmission by Vectors
Aphid vectors include: Amphorophora agathonica, Aphis rubifolii, Aulacorthum solani, Chaetosiphon fragaefolii, C. jacobi, C. tetrahodus, C. thomasi, Macrosiphum pelargonii and M. rosae, and Myzus ascalonicus, M. ornatus and M. persicae (Frazier, 1955; Mellor & Forbes, 1960). Of these, the most efficient are C. fragaefolii, C. jacobi and C. thomasi. Some strains of the virus are not transmitted by Chaetosiphon fragaefolii, C. thomasi or Myzus ornatus (Frazier, 1960; Mellor & Forbes, 1960). Aphids can acquire the virus during a 30 min acquisition access feeding period (Frazier, 1955). Retention of the virus by feeding vectors is variously reported as 1 h (Prentice, 1952), 8 h (Frazier, 1955) and 24 h (Schöniger, 1958) for various isolates of strawberry vein banding virus. In fasting vectors, a half-life of about 10 h was calculated by Frazier & Sylvester (1960). The virus was not retained through ecdysis by Chaetosiphon jacobi (Frazier, 1966).
Transmission through Seed
None reported.
Transmission by Dodder
Transmitted by Cuscuta subinclusa (Frazier, 1955).
Serology
No antiserum has been prepared. Partially purified virus reacted with globulin from cauliflower mosaic virus antiserum in enzyme-linked immunosorbent assay (ELISA) and immunospecific electron microscopy (ISEM) tests (Morris et al., 1980).
Relationships
1. Between strains. In cross-protection tests using seven isolates, complete or partial protection resulted whether the challenge virus was introduced by vectors or by grafting. Isolates from eastern North America appeared more closely related to each other than to western isolates (N. W. Frazier, unpublished data).
2. To other viruses. The virus reacts weakly with cauliflower mosaic virus antiserum; this, together with sedimentation behaviour and particle morphology, suggests that it is a distinct member of the caulimovirus group (Kitajima et al., 1973; Morris et al., 1980).
Stability in Sap
No information.
Purification
Morris et al. (1980). Homogenize infected leaves of Alpine strawberry (Fragaria vesca var. semperflorens ‘Alpine’) 1:5 (w:v) in 0.5 M potassium phosphate buffer, pH 7.5, plus 0.2% 2-mercaptoethanol; clarify by adding chloroform to 25% (v/v), and n-butanol to 7% (v/v) and centrifuging at low speed. Precipitate the virus from the supernatant fluid by adding NaCl to 0.1 M and polyethylene glycol (M. Wt 6000) to 8% (w/v). Resuspend the pellet in 100 ml distilled water with slow stirring overnight at 4°C. Float 15 ml of the preparation over 7 ml of 30% sucrose over a ‘cushion’ of 3 ml of 30% sucrose containing 1.5 g/cm3 CsCl in a Beckman SW 25.1 tube; centrifuge at 24,000 rev./min for 2 h at 4°C. The virus accumulates at the CsCl-sucrose interface, is observable there as an opalescent band, and can be directly detected by ELISA and ISEM with cauliflower mosaic virus antiserum globulin.
Properties of Particles
A sedimentation coefficient (s20,w) of 200 ± 10 S was calculated by co-sedimentation with cauliflower mosaic virus in rate-zonal sucrose density gradient centrifugation (Morris et al., 1980).
Particle Structure
Particles are isometric (Fig. 4) with diameter of 40-50 nm (Kitajima et al., 1973; Morris et al., 1980) or 43 ± 4 nm (R. H. Converse & J. L. Pyott, unpublished data). Details of ultrastructure are unknown.
Particle Composition
The particles probably contain double-stranded DNA, by analogy with other caulimoviruses.
Relations with Cells and Tissues
Virus particles occur in cytoplasmic inclusions in cells of the leaf vascular parenchyma immediately adjoining xylem vessels and, less frequently, in mesophyll and epidermal cells. The inclusions (Fig. 2, Fig. 3) are dense, usually with ellipsoidal profiles 1500 x 800 nm, but without a limiting outer membrane; they consist of a finely granular matrix, usually having several electron-transparent areas of varied size, and have ribosome-like granules attached to the outer surface. They resemble inclusions induced by other caulimoviruses (Kitajima et al., 1973; Morris et al., 1980; R. H. Converse & J. L. Pyott, unpublished data).
Notes
Severity of the strawberry vein banding disease varies widely with host, virus isolate and stage of infection. Infection is usually symptomless in Fragaria chiloensis and F. ananassa. If crinkle virus is also present, the indicator plants may show necrosis of young leaflets and petioles. Strawberry leaf curl (Prentice, 1952) and strawberry virus 5 (Prentice, 1952) are probably complexes of strawberry vein banding virus and the latent A strain of strawberry crinkle virus (Frazier, 1955). Strawberry vein banding virus can best be diagnosed by graft or aphid transmission to plants of the indicator clones Fragaria virginiana UC-12 or F. vesca UC-6 (Frazier, 1974b) which show characteristic chlorotic vein-banding. The virus can be separated from other viruses most readily by vector transmission. In chronically infected plants, the virus may become unavailable to the vectors (Prentice, 1952), but availability can be restored by graft transmission to a new plant (Frazier & Posnette, 1958). The virus can be eliminated from mother plants by tissue culture of 0.5-1.0 mm runner tips (Miller & Belkengren, 1963) or by heat treatment for 10 days at 42°C (Bolton, 1965). Stock certified to be free of the virus is commercially available.
Figures

Chlorotic veins of Fragaria vesca var. semperflorens cv. ‘Alpine’ infected by leaf grafting with the Frazier type isolate.

Vacuolate inclusion in xylem parenchyma of a leaf of Fragaria vesca cv. UC-6 graft inoculated with the Frazier type isolate. Bar represents 200 nm.
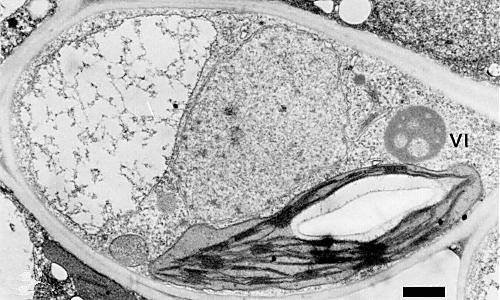
Vacuolate inclusion in xylem parenchyma cell of a leaf of Fragaria vesca var. semperflorens cv. ‘Alpine’ sampled 45 days after graft inoculation by an Oregon isolate. VI indicates virus inclusion. Bar represents 1 µm.
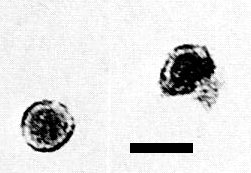
Particles in a partially purified preparation stained with uranyl acetate and detected by serologically specific electron microscopy using cauliflower mosaic virus antiserum. Bar represents 50 nm. (Courtesy T. J. Morris, Univ. of California, Berkeley.)
References list for DPV: Strawberry vein banding virus (219)
- Bolton, Res. Fmrs 10: 14, 1965.
- Frazier, Phytopathology 45: 307, 1955.
- Frazier, Pl. Dis. Reptr 44: 436, 1960.
- Frazier, Phytopathology 56: 64, 1966.
- Frazier, Pl. Dis. Reptr 58: 203, 1974a.
- Frazier, Pl. Dis. Reptr 58: 28, 1974b.
- Frazier & Posnette, Hilgardia 27: 455, 1958.
- Frazier & Sylvester, Virology 12: 233, 1960.
- Kitajima, Betti & Costa, J. gen. Virol. 20: 117, 1973.
- Mellor & Forbes, Can. J. Bot. 38: 343, 1960.
- Miller & Belkengren, Pl. Dis. Reptr 47: 298, 1963.
- Miller & Frazier, in Virus Diseases of Small Fruits and Grapevines, p. 8, ed. N. W. Frazier, Univ. of Calif. Div. Agric. Sci., Berkeley, Calif., 1970.
- Morris, Mullin, Schlegel, Cole & Alosi, Phytopathology 70: 156, 1980.
- Plakidas, Strawberry Diseases, p. 144, Louisiana State Univ. Press, Baton Rouge, 1964.
- Prentice, Ann. appl. Biol. 39: 487, 1952.
- Schöniger, Phytopath. Z. 32: 325, 1958.