Details of DPV and References
DPV NO: 222 September 1980
Family: Tymoviridae
Genus: Tymovirus
Species: Erysimum latent virus | Acronym: ErLV
Erysimum latent virus
D. D. Shukla Division of Protein Chemistry, CSIRO, Parkville (Melbourne), Victoria 3052, Australia
K. H. Gough Division of Protein Chemistry, CSIRO, Parkville (Melbourne), Victoria 3052, Australia
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
Described by
Shukla & Schmelzer (1972).
A virus with RNA-containing isometric particles c. 27 nm in diameter which sediment as two components. Its host range is confined almost entirely to the Cruciferae. The virus is readily sap-transmissible, and is transmitted in the field by flea-beetles. Occurs in the German Democratic Republic.
Main Diseases
Of the eight natural hosts found, Erysimum helveticum, E. perovskianum, E. pulchellum, E. sylvestre and Barbarea vulgaris were symptomlessly infected; mosaic symptoms in Arabis ludoviciana, Erysimum crepidifolium and Fibigia clypeata could have been caused by simultaneous infection with other viruses (Shukla, Proeseler & Schmelzer, 1975).
Geographical Distribution
Reported from several localities in the German Democratic Republic.
Host Range and Symptomatology
Readily transmitted by inoculation of sap to many species of Cruciferae and to a few members of the Cucurbitaceae, Labiatae, Leguminosae, Resedaceae and Solanaceae. Most infected plants show vein-clearing and mottling.
-
Diagnostic species
- Brassica chinensis
(pak-choi) and R. pekinensis (Chinese cabbage). Chlorotic or necrotic local lesions; plants showing chlorotic local lesions develop systemic vein-clearing, rings and mottling (Fig. 1); plants showing necrotic local lesions are not infected systemically. - R. juncea (Indian or Chinese mustard). Chlorotic and necrotic local
lesions, systemic
vein-clearing and mottling.
- R. napus var. napobrassica (swede). Numerous pin-point necrotic local lesions (Fig. 2) develop within a few days of inoculation; some plants show systemic mottling.
- Lunaria annua (honesty, satin flower). Systemic vein-clearing and mottling.
- Sinapis alba (white mustard). Chlorotic local lesions, systemic vein-clearing and mottling.
- R. napus var. napobrassica (swede). Numerous pin-point necrotic local lesions (Fig. 2) develop within a few days of inoculation; some plants show systemic mottling.
-
Propagation species
- Lunaria annua
is a good host for maintaining isolates. Brassica chinensis, B. juncea and Sinapis alba are good sources of virus particles for purification.Assay species
- Brassica napus
var. napobrassica is a very sensitive and reliable local lesion host (Shukla & Schmelzer, 1973a).
Strains
None found.
Transmission by Vectors
Transmitted by flea-beetles in four species of the genus Phyllotreta. The adult beetles transmitted the virus after acquisition access times of less than 10 min and inoculation access times of 30 to 60 min. After the acquisition feeding the insects remained infective for up to 3 days (Shukla et al., 1975; Proeseler & Schmelzer, 1977).
Transmission through Seed
Not seed-borne in Brassica chinensis.
Serology
Particles are strongly immunogenic in rabbits. Antisera with precipitin titres of 1/512-1/8192 have been produced.
Relationships
Serologically, the virus is related distantly to okra mosaic virus and very distantly to Andean potato latent and ononis yellow mosaic viruses. It has many properties typical of the tymovirus group including particle morphology and sedimentation behaviour, molecular weight of coat protein, high nucleic acid content of particles and high cytidine content of RNA, high thermal inactivation point, high concentration in host plants, beetle transmission and induction of double membrane-bounded vesicles in the chloroplasts of infected cells (Shukla et al., 1980).
Stability in Sap
In sap from infected Sinapis alba the virus loses infectivity after dilution between 10-5 and 10-6, heating for 10 min at between 76°C and 78°C or storage for 21 days at about 22°C.
Purification
(Shukla et al., 1973). Homogenize infected Brassica chinensis tissue in 0.05 M phosphate buffer, pH 7.0 (2 ml/g tissue). Clarify by adding an equal volume of a 1:1 mixture of n-butanol and chloroform. Concentrate and purify the particles by two cycles of differential centrifugation, resuspending the pellets in 0.05 M phosphate buffer, pH 7.0. Further purification may be achieved by sucrose density gradient centrifugation. Highest virus concentration in R. chinensis is achieved 2 weeks after inoculation when plants are grown at 22°C under continuous artificial light (Shukla & Schmelzer, 1973b; 1973c). Yields of 0.5 mg virus/g tissue are readily obtained. The virus crystallizes from 8% (w/v) polyethylene glycol, M. Wt 6000, either as truncated tetrahedra (Fig. 4) or as thin monoclinic plates (P. M. Colman, P. A. Tulloch, D. D. Shukla & K. H. Gough, unpublished results).
Properties of Particles
(K. H. Gough, G. G. Lilley, D. D. Shukla & E. F. Woods, unpublished results). The particles sediment as two components in sucrose gradients, a fast sedimenting (B) component of infective nucleoprotein particles, and a slower (T) component of protein shells (Fig. 5). The two components are serologically indistinguishable.
Sedimentation coefficients (s20,w) at infinite dilution (svedbergs): 59 (T), 113 (B).
Molecular weights (daltons): 3.9 x 106 (T), 5.9 x 106 (B).
Diffusion coefficient (D20 x 10-7 cm2/sec): 1.35 (B).
Isoelectric point: pH 5.40 ± 0.05 (B).
Partial specific volume: 0.680 (B).
Electrophoretic mobility: in immunoelectrophoresis in 1% agarose containing 0.025 M phosphate buffer, pH 7.0, the virus migrates slowly towards the anode.
Absorbance at 260 nm (1 mg/ml, 1 cm light path): 9.7 (B).
A260/A280: 1.76 (B).
Buoyant densities in CsCl (g/cm3): 1 .26 (T), 1 .42 (B).
Particle Structure
Particles are about 27 nm in diameter and have icosahedral symmetry (Fig. 3). They contain 180 protein subunits clustered into 20 hexamers and 12 pentamers. Electron micrographs of negatively stained virus show 32 morphological subunits (P. M. Colman, P. A. Tulloch, D. D. Shukla & K. H. Gough, unpublished results).
Particle Composition
Nucleic acid: RNA, single-stranded, c. 32% of particle weight. M. Wt, estimated from particle composition and structure, c. 2.0 x 106. Nucleotide composition G 15.6: A 23.9: C 34.5; U 26.0 (Shukla et al., 1980). RNA extracted with phenol had about 14% of the infectivity expected from that of the intact virus (Shukla et al., 1973).
Protein: 68% of particle by weight. Single species of M. Wt 21,700, determined by electrophoresis in polyacrylamide gels containing sodium dodecyl sulphate. Ala 14 residues, arg 5, asp 11, cys 0, glu 21, gly 17, his 4, ile 8, leu 22, lys 7, met 3, phe 7, pro 21, ser 28, thr 20, trp 1, tyr 3, val 16; total 208 (Shukla et al., 1980).
Relations with Cells and Tissues
The virus, like other tymoviruses, induces the formation of small vesicles, bounded by double membranes, at the peripheries of the chloroplasts in infected cells, e.g. in Chinese cabbage and turnip (Fig. 6). Many of the chloroplasts are rounded but few show the vacuolation induced by other tymoviruses. The nuclei contain large masses of lightly staining virus protein shells. The cytoplasm contains many darkly staining virus particles (Shukla et al., 1980).
Notes
Radish mosaic, turnip crinkle, turnip rosette, turnip yellow mosaic and erysimum latent are all beetle-transmitted viruses with isometric particles about 30 nm in diameter, isolated from species of the Cruciferae. Host reaction and host range cannot reliably distinguish among them. The five viruses are unrelated to each other serologically. Erysimum latent and turnip yellow mosaic viruses possess similar biophysical properties (tymoviruses) which distinguish them from the other three viruses.
Figures

Vein-clearing and mottling in systemically infected leaf of Brassica chinensis.

Pin-point local lesions in inoculated leaf of Brassica napus var. napobrassica.

Virus particles from a purified preparation, negatively stained with ammonium molybdate. Bar represents 100 nm. (Photo courtesy of P. Tulloch.)

Truncated tetrahedral crystals formed in 8% (w/v) polyethylene glycol, M. Wt 6000. Bar represents 1 mm.
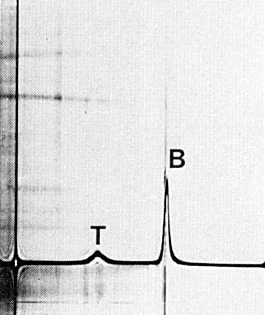
Schlieren diagram obtained by analytical ultracentrifugation of a purified virus preparation showing top (T) and bottom (B) components. Sedimentation from left to right.
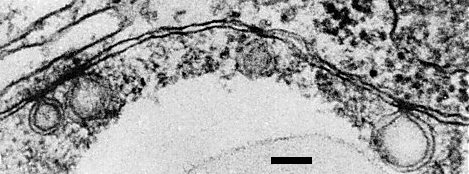
Ultrathin section of infected turnip leaf showing vesicles bounded by double membranes at the periphery of a chloroplast. Bar represents 100 nm.
References list for DPV: Erysimum latent virus (222)
- Proeseler & Schmelzer, Zentbl. Bakt. ParasitKde, Abt. 2 132: 716, 1977.
- Shukla & Schmelzer, Acta phytopath. Acad. Sci. hung. 7: 157, 1972.
- Shukla & Schmelzer, Acta phytopath. Acad. Sci. hung. 8: 105, 1973a.
- Shukla & Schmelzer, Acta phytopath. Acad. Sci. hung. 8: 113, 1973b.
- Shukla & Schmelzer, Acta phytopath. Acad. Sci. hung. 8: 121, 1973c.
- Shukla, Proll, Schmelzer & Schmidt, Acta phytopath. Acad. Sci. hung. 8: 91, 1973.
- Shukla, Proeseler & Schmelzer, Acta phytopath. Acad. Sci. hung. 10: 211, 1975.
- Shukla, Koenig, Gough, Huth & Lesemann, Phytopathology 70: 382, 1980.