Details of DPV and References
DPV NO: 223 September 1980
Family: Virgaviridae
Genus: Pomovirus
Species: Broad bean necrosis virus | Acronym: BBNV
Broad bean necrosis virus
T. Inouye College of Agriculture, University of Osaka Prefecture, Sakai, Osaka, Japan
W. Nakasone Osaka Agricultural Research Centre, Habikino, Osaka, Japan
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
Disease described by
Fukano & Yokoyama (1951),
and virus partially characterized by
Inouye & Asatani (1968).
A virus with straight rod-shaped particles of two lengths, c. 150 and 250 nm, and both c. 25 nm wide. It has a somewhat limited host range and is transmitted by mechanical inoculation of sap and through soil. Found in Japan.
Main Diseases
The virus is found in nature only in broad bean; the leaves develop reddish brown spots, rings, veinal necrosis and streaks (Fig. 1, Fig. 2). The virus often produces chlorotic spots along the veins of expanding young leaves. Severely affected plants are stunted and defoliated. Symptoms usually appear in early spring but tend to disappear as the weather becomes warmer.
Geographical Distribution
Found in Japan.
Host Range and Symptomatology
The virus infected 10 species in 4 dicotyledonous families out of 35 species in 9 families tested (Inouye & Asatani, 1968; T. Inouye, unpublished data). Broad bean (Vicia faba), pea (Pisum sativum) and sweet pea (Lathyrus odoratus ) are infected systemically. Symptoms are not produced in the glasshouse at temperatures much above 20°C.
-
Diagnostic species
- Vicia faba
(broad bean). Reddish brown necrotic local lesions develop in inoculated leaves; the systemic symptoms are necrotic spots, rings, veinal necrosis in the leaves (Fig. 1, Fig. 2) and streaks on stems and petioles. Chlorotic spots often occur along the veins of newly unfolded leaves and later the tissue around the spots usually becomes necrotic. -
Pisum sativum (pea). Mild systemic mottling, often accompanied by necrotic flecks, veinal
necrosis and streaks. Severely affected plants are stunted.
- Phaseolus vulgaris (bean) cv. Yamashiro Kurosando. Necrotic rings in inoculated leaves (Fig. 3); no systemic infection.
- Nicotiana clevelandii. Chlorotic local lesions (Fig. 6); no systemic infection.
- Chenopodium amaranticolor. Local chlorotic and semi-necrotic concentric rings (Fig. 4); no systemic infection.
- C. quinoa. Local necrotic spots or rings; no systemic infection.
- Phaseolus vulgaris (bean) cv. Yamashiro Kurosando. Necrotic rings in inoculated leaves (Fig. 3); no systemic infection.
-
Propagation species
- Pea (Pisum sativum) and sweet pea (Lathyrus odoratus) can be used for maintaining cultures and for propagating the virus.
-
Assay species
- Chenopodium amaranticolor
and C. quinoa can be used for local lesion assay.
Strains
No strains have been reported.
Transmission by Vectors
Infectivity is retained for more than 1 year in air-dried soil (Fujikawa, 1963) suggesting that the virus has a soil-inhabiting fungal vector. Several phycomycetous fungi, including Polymyxa and Olpidium, are found in the roots of broad beans grown in infective soil, but no species has been proved to be a vector (W. Nakasone & T. Inouye, unpublished observations).
Transmission through Seed
Not found in broad bean (Fujikawa, 1963).
Serology
The virus is moderately immunogenic: antiserum from a rabbit given four intramuscular injections of the virus emulsified in Freund’s complete adjuvant had a homologous titre in microprecipitin tests of 1/512 (Nakasone & Inouye, 1977).
Relationships
The virus is classified in the tobamovirus group on the basis of particle morphology and serological relationships to other tobamoviruses. Microprecipitin tests and electron microscope serology indicate that the virus is distantly serologically related to cucumber green mottle mosaic, odontoglossum ringspot and soil-borne wheat mosaic viruses (Nakasone & Inouye, 1978). No serological relationships were detected to tobacco mosaic virus (Nakasone & Inouye, 1978) or hypochoeris mosaic virus (Brunt & Stace-Smith, 1978).
Stability in Sap
In pea sap, the thermal inactivation point (10 min) is 55 to 60°C, and the dilution end-point is between 10-3 and 10-4. Infectivity is retained for 8 days at 20°C (Inouye & Asatani, 1968).
Purification
Partially purified virus was obtained by the following procedures (Nakasone & Inouye, 1977). Homogenize dried tissues of diseased pea in 0.1 M citrate buffer (pH 7.5) containing 0.1% ascorbic acid and 1% Triton X-100. Clarify expressed juice by adding 0.5 vol. ethyl ether to 1 vol. extract. Precipitate the virus by adding polyethylene glycol, M. Wt 6000 (PEG), to 4% (w/v). Centrifuge at low speed and resuspend the precipitate in 0.05 M citrate buffer (pH 7.5); then clarify further by adding 0.25 vol. ethyl ether to 1 vol. extract. Precipitate the virus by adding PEG to 4% (w/v) and NaCl to 2% (w/v) and resuspend in 0.05 M citrate buffer (pH 7.5). Sediment the virus by high speed centrifugation through a 20% sucrose cushion, and resuspend the pellet in distilled water.
Properties of Particles
Not known.
Particle Structure
Particles are straight rods (Fig. 5), c. 25 nm wide, with two lengths, c. 150 and 250 nm (Inouye & Asatani, 1968). Negatively stained particles show helical symmetry.
Particle Composition
No information.
Relations with Cells and Tissues
Amorphous cellular inclusion bodies can be found by light microscopy in cells of epidermal strips of infected leaves of broad bean, pea and sweet pea. Examination of ultrathin sections of infected leaf tissues of broad bean and sweet pea in the electron microscope reveals prominent membranous network structures often associated with lipid-like globules. Virus particles occur in the cytoplasm either separately or in loose aggregates (Fig. 7) (Inouye, 1971).
Notes
Broad bean necrosis virus can be distinguished from other elongated viruses that infect broad beans in Japan by particle morphology and mode of transmission. Bean yellow mosaic virus and pea seed-borne mosaic virus have flexuous elongated particles and are transmitted by aphids and through seed, whereas broad bean necrosis virus has straight rod-shaped particles and is transmitted through the soil. In some respects, broad bean necrosis virus resembles other viruses having straight rod-shaped particles, especially the fungus-transmitted tobamoviruses such as beet necrotic yellow vein (Tamada, 1975), potato mop-top (Harrison, 1974) and soil-borne wheat mosaic viruses (Brakke, 1971). However, in host range and symptomatology it is clearly distinct from these viruses and from three other possible members of the tobamovirus group, hypochoeris mosaic (Brunt & Stace-Smith, 1978), Nicotiana velutina mosaic (Randles, 1978) and peanut clump (Thouvenel, Dollet & Fauquet, 1976) viruses. The last virus can infect leguminous plants, but is clearly distinct from broad bean necrosis virus in its particle dimensions, host range and optimal temperatures for multiplication (Thouvenel et al., 1976).
Figures

Systemic necrotic spots, rings and veinal necrosis in Vicia faba.

Necrotic symptoms in naturally infected V. faba leaf.

Necrotic lesions in inoculated leaf of Phaseolus vulgaris cv. Yamashiro Kurosando.

Concentric ring lesions in inoculated leaf of Chenopodium amaranticolor.
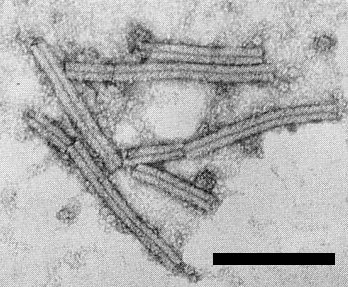
Virus particles stained with uranyl formate. Bar represents 200 nm.

Chlorotic lesions in inoculated leaf of Nicotiana clevelandii.

Section of portion of a cell from a systemically infected leaf of V. faba, showing prominent membranous structures, lipid-like globules and loosely aggregated virus particles in the cytoplasm. Bar represents 500 nm.
References list for DPV: Broad bean necrosis virus (223)
- Brakke, CMI/AAB Descriptions of Plant Viruses 77, 4 pp., 1971.
- Brunt & Stace-Smith, Ann. appl. Biol. 90: 205, 1978.
- Fujikawa, Nogyo oyobi Engei 38: 73, 1963.
- Fukano & Yokoyama, Kyushu Nogyo Kenkyu 10: 133, 1951.
- Harrison, CMI/AAB Descriptions of Plant Viruses 138, 4 pp., 1974.
- Inouye, Ber. Ohara Inst. landw. Biol. 15: 69, 1971.
- Inouye & Asatani, Ann. phytopath. Soc. Japan 34: 317, 1968.
- Nakasone & Inouye, Ann. phytopath. Soc. Japan 43: 100, 1977.
- Nakasone & Inouye, Ann. phytopath. Soc. Japan 44: 97, 1978.
- Randles, CMI/AAB Descriptions of Plant Viruses 189, 4 pp., 1978.
- Tamada, CMI/AAB Descriptions of Plant Viruses 144, 4 pp., 1975.
- Thouvenel, Dollet & Fauquet, Ann. appl. Biol. 84: 311, 1976.