Details of DPV and References
DPV NO: 227 September 1980
Family: Tombusviridae
Genus: Carmovirus
Species: Hibiscus chlorotic ringspot virus | Acronym: HCRSV
Hibiscus chlorotic ringspot virus
H. Waterworth U.S. Department of Agriculture, Science & Education Administration, Plant Introduction Station, Glenn Dale, Maryland 20769, USA
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
Described by
Waterworth, Lawson & Monroe (1976).
A virus with RNA-containing isometric particles c. 28 nm in diameter sedimenting as a single component. It is readily mechanically transmitted, infecting species primarily in the Malvaceae, but is probably of limited economic importance in this family. It exists throughout the world.
Main Diseases
Occurs naturally in many Hibiscus rosa-sinensis cultivars (Jones & Behncken, 1980), in which it incites a generalized mottle (Fig. 1), or numerous tiny chlorotic spots (Fig. 2), rings (Fig. 3, Fig. 8) or vein-banding on most leaves of infected plants. Plants continue to flower and are not noticeably stunted.
Geographical Distribution
Australia, USA, Fiji, El Salvador, Thailand.
Host Range and Symptomatology
Incites systemic mosaics in species of Malvaceae. A few members of Amaranthaceae, Chenopodiaceae, Leguminosae and Scrophulariaceae respond with local lesions only. It did not infect the plants tested in 20 other families (Waterworth et al., 1976). Readily transmitted by manual inoculation of sap.
-
Diagnostic species
- Chenopodium quinoa.
Chlorotic local lesions, 2-3 mm in diameter in 10 days; no systemic infection. - Cyamopsis tetragonoloba (guar). Distinct 0.5 mm diameter necrotic local lesions
(Fig. 4)
in 6 days; no systemic infection.
- Dolichos biflorus. Distinct 1 mm diameter necrotic local lesions (Fig. 9) in 6 days; no systemic infection.
- Hibiscus cannabinus (kenaf). Local lesions in 6 days (Fig. 5) and systemic mottle (Fig. 6) in 10 to 20 days.
- Gossypium hirsutum (cotton). Distinct 0.5 mm diameter necrotic local lesions (Fig. 7) in 15 to 20 days; no systemic infection.
- Phaseolus vulgaris cv. 'Pinto' (French bean). Pin-point local lesions in 6 days; no systemic symptoms.
- Dolichos biflorus. Distinct 1 mm diameter necrotic local lesions (Fig. 9) in 6 days; no systemic infection.
-
Propagation species
- Hibiscus cannabinus
is an acceptable source of virus for purification. Though yields of tissue are low, the plant grows quickly, is easily infected systemically, and can be harvested over a long period of time. Althaea spp. and H. rosa-sinensis (Chinese hibiscus) are suitable species for maintaining cultures.Assay species
- Cyamopsis tetragonoloba
and Dolichos biflorus are suitable for local lesion assay (Fig. 4, Fig. 9). Hibiscus cannabinus is the most sensitive host; some inocula that fail to incite symptoms in other assay species may incite distinct mottle (Fig. 6) in this species although the symptoms may take 2 to 3 weeks to appear.
Strains
None reported.
Transmission by Vectors
Not transmitted by the aphid Myzus persicae after short acquisition feeds, nor by the Mexican bean beetle, Epilachna varivestis.
Transmission through Seed
No information.
Serology
The virus is a good immunogen; antiserum titres of 1/1024 are readily attainable. In the droplet microprecipitin test partially purified preparations of the virus give a distinct reaction within 30 min. Double diffusion in gels containing 0.75% agar and 0.02% sodium azide in water, is an excellent procedure for identifying the virus in crude extracts of most herbaceous species but does not detect the virus in infected leaves of the woody H. rosa-sinensis. A single, sharp, curved band can be observed within 24 h with most virus sources.
Relationships
The virus is serologically unrelated to any of 45 other viruses tested with spherical particles (Waterworth et al., 1976), including okra mosaic virus (Givord & Hirth, 1973) and hibiscus latent ringspot virus (Brunt et al., 1980). Isolates from the USA and El Salvador (Waterworth et al., 1976), Australia (Jones & Behncken, 1980), Fiji (A. A. Brunt, unpublished data) and Thailand (H. C. Phatak, unpublished data) were serologically indistinguishable. Its relationship to hibiscus ringspot virus (Lana, 1974) is unknown.
Stability in Sap
In kenaf sap, the thermal inactivation point (10 min) is about 72°C, dilution endpoint about 10-8, and infectivity is retained at 20°C for about 30 days. In these tests the time required for systemic symptoms to appear in kenaf plants in the glasshouse ranged from 14 to 49 days and was inversely related to the concentration of infective virus (Waterworth et al., 1976).
Purification
Blend each 100 g fresh kenaf leaves in 300 ml 0.01 M phosphate-borate buffer (Waterworth et al., 1976) and then add 20 ml chloroform. Centrifuge at 12,000 g for 10 min. Concentrate the virus from the clear yellow supernatant fluid by one of three methods: (1) centrifuge at 100,000 g for 75 min, (2) mix with equal volume of a saturated solution of ammonium sulphate and centrifuge at low speed, or (3) add polyethylene glycol (M. Wt 6000) to 10% (w/v), without NaCl, and centrifuge at low speed. Resuspend all pellets in 0.2 M sodium acetate buffer (pH 6.0) and centrifuge at 10,000 g for 5 min. Further purify in 10 to 40% sucrose gradients. Dialyse virus fractions, concentrate the virus by high speed centrifugation and store in acetate buffer. Yields of virus are 0.3 to 0.5 mg/g tissue.
Properties of Particles
Single component (Fig. 10) with sedimentation coefficient (s20,w) about 118 S (Jones & Behncken, 1980; Waterworth et al., 1976; R. Hull, unpublished data).
A260/A280: 1.49 (Waterworth et al., 1976; Jones & Behncken, 1980);
Amax(260)/Amin(242): 1.19, both uncorrected for light-scattering.
Absorbance at 260 nm (1 mg/ml, 1 cm light path): 4.6 (Waterworth et al., 1976).
Particle Structure
Particles are isometric, about 28 nm in diameter. Negative stain apparently penetrates about one particle in five (Fig. 11) (Waterworth et al., 1976; Jones & Behncken, 1980).
Particle Composition
Nucleic acid: RNA, single-stranded, M. Wt about 1.55 x 106, comprising about 14% of particle weight.
Molar percentages of nucleotides: G:23.8; A:25.5; C:26.4; U:24.3. The N:P ratio is about 11.2 (Waterworth et al., 1976).
Protein: Not determined.
Other components: None reported.
Relations with Cells and Tissues
Virus particles are scattered in the cytoplasm and are not in crystalline arrays. They occur in clusters in the central vacuoles in some cells. Clusters may form more readily in cells in which the tonoplast has been disrupted (R. H. Lawson, unpublished data).
Notes
Symptoms associated with hibiscus chlorotic ringspot virus in H. rosa-sinensis vary considerably. This may be attributable to such variables as age of the plants or leaves, or to environmental factors, but may also be caused by genetic variability within the species. For example, leaf edges in some cultivars are distinctly serrated (Fig. 1) whereas those of others are entire (Fig. 2). Leaf shape varies even on a single plant. The virus was named primarily on the basis of the symptoms incited upon mechanical inoculation of healthy seedlings (Fig. 10).
Hibiscus chlorotic ringspot virus is probably not the cause of most of the virus-like diseases reported in malvaceous plants. For example, it is not related to the tymovirus that causes okra mosaic (okra = Hibiscus esculentus) (Givord & Hirth, 1973); it showed no relationship in serological tests to the three component hibiscus latent ringspot virus (Brunt et al., 1980) which is associated with a mild leaf chlorosis of H. rosa-sinensis; and it is not related to malva yellows virus (Costa et al., 1959), an aphid-transmissible virus that exists naturally in many species that hibiscus chlorotic ringspot virus does not infect. Moreover, it is probably not related to the various whitefly-transmissible agents that cause chloroses of species of Malvaceae (Flores & Silberschmidt, 1967; Costa, 1954), and yellow vein mosaics of bhendi (Hibiscus esculentus) (Capoor & Varma, 1950), okra and cotton (Sastry & Singh, 1973). Likewise it seems to be unrelated to the non-mechanically transmitted agents of hibiscus line pattern (Wolfswinkel, 1966) and leaf curl (Mukherjee & Raychaudhuri, 1964) because the symptoms are very different. Except for okra mosaic virus and hibiscus latent ringspot viruses, attempts to transmit the above agents mechanically have failed.
Hibiscus chlorotic ringspot virus may be related to or identical with the agents that cause a mosaic disease of H. rosa-sinensis in Hawaii (Hendrix & Murakishi, 1951), a mosaic in H. manihot in New Guinea (Van Velsen, 1967), and a hibiscus ringspot disease in Nigeria (Lana, 1974). Viruses described in the two last-named reports were mechanically transmitted.
Figures

Leaf of infected Hibiscus rosa-sinensis (Chinese hibiscus) from Queensland. (Courtesy Jones & Behncken, 1980.)

Leaf of naturally infected Hibiscus rosa-sinensis from the USA.

Leaf of naturally infected Hibiscus sp. from Thailand.
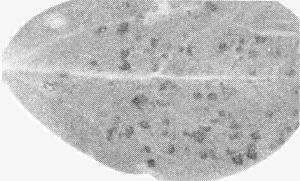
Necrotic local lesions in Cyamopsis tetragonoloba (guar) leaf.

Necrotic local lesions in H. cannabinus (kenaf) leaf.

Systemic mosaic in kenaf leaf.

Necrotic local lesions in Gossypium hirsutum (cotton) leaf.
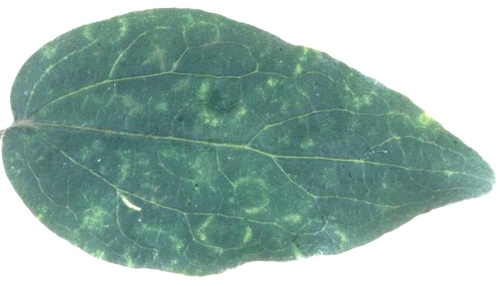
Systemic chlorotic ringspots in leaf of H. rosa-sinensis mechanically inoculated with purified virus.

Necrotic local lesions in Dolichos biflorus leaf.

Schlieren diagram of moving boundary analytical ultracentrifugation of purified virus in 0.02 M sodium acetate buffer, pH 6; photograph taken 12 min after attaining speed of 35,600 rev./min. (Courtesy R. Hull.)

Particles from a purified preparation stained with potassium phosphotungstate, pH 6.8. About 20% of the particles are disintegrated or are penetrated by the stain. Bar represents 100 nm. (Courtesy R. Lawson.)
References list for DPV: Hibiscus chlorotic ringspot virus (227)
- Brunt, Barton, Phillips & Lana, Ann. appl. Biol. 96: 37, 1980.
- Capoor & Varma, Ind. J. agric. Sci. 20: 217, 1950.
- Costa, Bragantia 13: 23, 1954.
- Costa, Duffus & Bardin, J. Am. Soc. Sug. Beet Technol. 10: 371, 1959.
- Flores & Silberschmidt, Phytopath Z. 60: 181, 1967.
- Givord & Hirth, Ann. appl. Biol. 74: 359, 1973.
- Hendrix & Murakishi, Bienn. Rep. Hawaii Univ. agric. Exp. Stn 1948-1950: 123, 1951.
- Jones & Behncken, Australas. Pl. Path. 9: 4, 1980.
- Lana, Pl. Dis. Reptr 58: 1040, 1974.
- Mukherjee & Raychaudhuri, Indian J. Hort. 21: 176, 1964.
- Sastry & Singh, Indian Phytopath. 26: 136, 1973.
- van Velsen, Papua New Guin. agric. J. 19: 10, 1967.
- Waterworth, Lawson & Monroe, Phytopathology 66: 570, 1976.
- Wolfswinkel, S. Afr. J. agric. Sci. 9: 483, 1966.