Details of DPV and References
DPV NO: 228 September 1980
Family: Betaflexiviridae
Genus: Vitivirus
Species: Heracleum latent virus | Acronym: HLV
Heracleum latent virus
F. Bem Benaki Phytopathological Institute, Kiphissia, Athens, Greece
A. F. Murant Scottish Horticultural Research Institute, Invergowrie, Dundee, Scotland
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
Described by
Murant & Goold (1972) and
Bem & Murant (1979a,
1979b,
1979c).
Synonym
- HV2 (Murant & Goold, 1972)
-
A virus with very flexuous filamentous RNA-containing particles c. 730 x 12 nm. Occurs symptomlessly in Heracleum sphondylium from which it is transmissible by aphids (Cavariella spp.) in the semi-persistent manner. Transmissible by inoculation of sap to a moderate range of herbaceous species.
Main Diseases
Occurs in hogweed (Heracleum sphondylium; Umbelliferae) without causing obvious symptoms. Not reported from cultivated umbelliferous plants.
Geographical Distribution
Reported from the UK. Also found in the Netherlands (L. Bos, personal communication).
Host Range and Symptomatology
The virus is readily transmitted by inoculation of sap; it infected 39 species from 11 families of dicotyledonous plants, especially species in the Amaranthaceae, Chenopodiaceae, Solanaceae and Umbelliferae (Bem & Murant, 1979b). Most species were infected symptomlessly.
-
Diagnostic species
- Chenopodium quinoa.
Necrotic local lesions in 6-7 days, downward reflexing of the petioles of inoculated leaves, followed 4-5 days later by characteristic systemic epinasty and stunting (Fig. 2, Fig. 3). - C. amaranticolor. Slight curling and vein-clearing of systemically infected
leaves. In winter, inoculated leaves show small chlorotic spots 1 mm in diameter after
7-8 days.
- Coriandrum sativum (coriander). Symptomless systemic infection or mild interveinal yellowing of systemically infected leaves.
- Nicotiana clevelandii, N. tabacum (cvs Samsun and White Burley) and Cyamopsis tetragonoloba (guar) are not infected.
- Coriandrum sativum (coriander). Symptomless systemic infection or mild interveinal yellowing of systemically infected leaves.
-
Propagation species
- Chenopodium quinoa
is suitable for maintaining cultures and is a good source of virus for purification.Assay species
- Chenopodium murale
(Fig. 1) is a useful local-lesion host. Coriandrum sativum is a useful test plant for aphid-transmission tests but the presence of the virus must be confirmed by mechanical inoculation of sap to C. quinoa.
Strains
No strains have been distinguished.
Transmission by Vectors
Transmitted in the semi-persistent manner by the aphids Cavariella aegopodii, C. pastinacae and C. theobaldi. Aphids transmitted the virus from naturally infected hogweed plants but not from those inoculated manually nor from aphid-inoculated plants of carrot (Daucus carota), celery (Apium graveolens), chervil (Anthriscus cerefolium), coriander (Coriandrum sativum), Cryptotaenia japonica, dill (Anethum graveolens) or parsley (Petroselinum crispum). Transmission by aphids may therefore depend on a helper virus present in naturally infected Heracleum sphondylium but not able to infect other umbelliferous species so far tested (A. F. Murant & F. Bem, unpublished data). Transmission was not assisted by anthriscus yellows virus, which assists transmission of parsnip yellow fleck virus (Murant & Goold, 1968).
Aphids can acquire the virus after acquisition access times of more than 2 h, the efficiency of transmission increasing with longer acquisition access times. Infective adult aphids retained the ability to inoculate the virus for 1 or 2 days, but not 3 days, after removal from the source plant (A. F. Murant & F. Bem, unpublished data).
Transmission through Seed
Not transmitted through the seed of C. quinoa, chervil, coriander or hogweed. For each species, 200 seedlings were tested (Bem & Murant, 1979b).
Serology
Moderately immunogenic. A rabbit given two intramuscular injections, one month apart, with formaldehyde-treated virus preparations gave a specific antiserum, 15 days after the second injection, with a titre of 1/1024 (Bem & Murant, 1979b). Microprecipitin tests can be done using purified or partially purified virus preparations. Specific precipitates of virus in serological tests in liquids are of the flocculent (flagellar) type. Latex flocculation tests give specific positive results with crude buffered extracts from infected C. quinoa, coriander, carrot or hogweed.
Relationships
Heracleum latent virus resembles the tentative closterovirus, apple chlorotic leaf spot, in particle morphology (very flexuous filaments 730 nm long with obvious cross-banding) and in physical and chemical properties (M. Wt of RNA and coat protein, sedimentation coefficient, particle stability, high A260/A280 ratio and lack of tryptophan). It also resembles definitive closteroviruses in many of these properties and in being transmitted by aphids in the semi-persistent manner. Thus it should probably be classed as a tentative closterovirus. However, it did not react with antisera to apple or prunus strains of apple chorotic leaf spot virus, or to six definitive closteroviruses (beet yellows, beet yellow stunt, carnation necrotic fleck, citrus tristeza, lilac chlorotic leaf spot and wheat yellow leaf viruses). It also failed to react with antiserum to apple stem grooving, potato T, wineberry latent, celery mosaic or parsnip mosaic viruses, or to several other viruses with filamentous particles belonging to the potyvirus, potexvirus and carlavirus groups (Bem & Murant, 1979b). Cross-protection tests in C. quinoa also gave no evidence of relationship to apple chlorotic leaf spot or apple stem grooving viruses (Bem & Murant, 1979b).
Stability in Sap
In bentonite-clarified extracts from C. quinoa, the thermal inactivation point (10 min) was 40-50°C, the dilution end-point was 10-4-10-5, and infectivity was abolished in 1-2 days at 22°C but survived for more than 6 days at 4°C. Infectivity was not stabilised by addition of reducing agents, divalent cations (Mg2+, Ca2+, Ni2+) or polyamines to leaf extracts made in 0.01 M acetate, pH 5.2, and was abolished by addition of ethylenediamine-tetraacetate (Bem & Murant, 1979b).
Purification
The following method was used by Bem & Murant (1979b): Do all steps at 4°C. Blend each 100 g infected C. quinoa tissue (whole plants) in 300 ml 0.01 M acetate buffer, pH 5.0. Squeeze the pulp through cheesecloth and re-extract the fibre with 100 ml of the above buffer. Combine the extracts and clarify by cautiously adding bentonite suspension (prepared according to Lister & Hadidi, 1971) in steps, starting with an initial amount of 9 ml bentonite suspension per 100 g tissue extracted. Centrifuge at 8000 g for 5 min. Precipitate the virus from the clarified extracts by adding polyethylene glycol (M. Wt 6000) to 8% (w/v) and allow to stand for 1 h. Centrifuge at 10,000 g for 20 min and resuspend the virus in 0.01 M Tris-HCl buffer, pH 7.6. Further purify and concentrate the virus preparations by one cycle of differential centrifugation followed by centrifugation in 10-40% sucrose density gradients. Yields are erratic, averaging c. 5 mg virus per 100 g tissue, but may reach up to 24 mg/100 g tissue.
Properties of Particles
(Bem & Murant, 1979c). Purified preparations form a single light-scattering band on sedimentation in sucrose density gradients.
Sedimentation coefficient (s°20,w) about 96 S (Fig. 4).
Buoyant density in Cs2SO4: 1.24 g/cm3.
A260/A280: about 1.5. Prolonged dialysis against 0.01 M Tris-HCl buffer, pH 7.6, does not alter this ratio.
Particle Structure
Particles are very flexuous filaments (Fig. 5) of modal length (in 2% phosphotungstate, pH 7) c. 730 nm, width 12.5 nm, and with obvious cross-banding (Fig. 6) of pitch c. 3.8 nm. The best stain for electron microscopy is 2% phosphotungstate, pH 7; breakages occur in ammonium molybdate (Bem & Murant, 1979b).
Particle Composition
Nucleic acid: Single-stranded RNA, M. Wt about 2.3 x 106 daltons, estimated by polyacrylamide gel electrophoresis under non-denaturing conditions (Bem & Murant, 1979c), or about 2.5 x 106 daltons, estimated by electrophoresis of glyoxal-denatured RNA in agarose gels (Murant, Taylor & Duncan, 1979).
Protein: Subunits have M. Wt about 23,500 daltons, estimated by SDS/polyacrylamide gel electrophoresis. The protein apparently lacks tryptophan (Bem & Murant, 1979c).
Other components: Does not contain carbohydrate (Bem & Murant, 1979c).
Relations with Cells and Tissues
All tissues are infected but the virus occurs in highest concentration in midrib, petiole and stem. Ultrathin sections of petioles or veins of C. quinoa, chervil and coriander reveal large aggregates in the phloem parenchyma cells and sieve tubes, composed of a fibrous material which is too thin to be P-protein and might represent aggregated virus particles, although this is not certain (F. Bem, A. F. Murant & I. M. Roberts, unpublished data).
Notes
Heracleum latent virus is common in hogweed, often in mixed infections with other viruses (Bem & Murant, 1979a) but its economic importance is unknown. Experimentally it infected several cultivated umbellifers, including carrot, celery, parsley and parsnip. It is readily distinguished from other viruses found in hogweed by the characteristic systemic epinasty it induces in C. quinoa and also by its systemic infection of C. amaranticolor, its mild symptoms in coriander, and its failure to infect Cyamopsis tetragonoloba and Nicotiana clevelandii (Bem & Murant, 1979a). Another closterovirus occurring in hogweed in UK and code-named HV6 (Bem & Murant, 1979a) has particles c. 1400 nm long and is not transmissible manually.
Figures

Small necrotic local lesions in Chenopodium murale 7 days after inoculation.

Necrotic local lesions in C. quinoa 7 days after inoculation.

A plant of C. quinoa 15 days after inoculation showing local lesions, downward reflexing of the petioles of inoculated leaves, and crinkling and epinasty of systemically infected leaves.
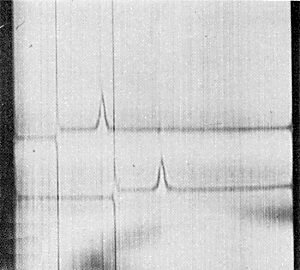
Schlieren pattern produced by (above) heracleum latent virus, (below) apple chlorotic leaf spot virus after 6 min at 30,000 rev./min. Sedimentation is from left to right.

Particles from sap of infected C. quinoa on antiserum-coated grids. Stained with 2% potassium phosphotungstate, pH 7. Bar represents 200 nm.
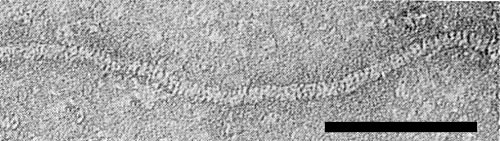
Portion of a particle stained with uranyl formate/NaOH, pH 4.8, showing cross-banding. Bar represents 100 nm.
References list for DPV: Heracleum latent virus (228)
- Bem & Murant, Ann. appl. Biol. 92: 237, 1979a.
- Bem & Murant, Ann. appl. Biol. 92: 243, 1979b.
- Bem & Murant, J. gen. Virol. 44: 817, 1979c.
- Lister & Hadidi, Virology 45: 240, 1971.
- Murant & Goold, Ann. appl. Biol. 62: 123, 1968.
- Murant & Goold, Rep. Scott. hort. Res. Inst. for 1971: 63, 1972.
- Murant, Taylor & Duncan, Rep. Scott. hort. Res. Inst. for 1978: 98, 1979.