Details of DPV and References
DPV NO: 233 July 1981
Family: Secoviridae
Genus: Nepovirus
Species: Hibiscus latent ringspot virus | Acronym: HLRSV
Hibiscus latent ringspot virus
A. A. Brunt Glasshouse Crops Research Institute, Littlehampton, Sussex, England
R. J. Barton Glasshouse Crops Research Institute, Littlehampton, Sussex, England
Sue Phillips Glasshouse Crops Research Institute, Littlehampton, Sussex, England
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
Described by Brunt et al. (1980).
A virus with isometric particles about 28 nm in diameter which sediment as three components and contain either no RNA or one or other of two species of single-stranded RNA. The virus is readily sap-transmissible to a moderately wide range of hosts, but its vector is unknown. Reported only from Nigeria.
Main Diseases
Hibiscus (Hibiscus rosa-sinensis) is the only known natural host. Some leaves of infected plants are faintly chlorotic but most, like the flowers, are symptomless.
Geographical Distribution
Reported only from Nigeria.
Host Range and Symptomatology
Transmitted readily by mechanical inoculation of sap to 22 of 74 species from 7 of 20 families (Brunt et al., 1980).
- Diagnostic species
- Chenopodium murale. Numerous fawn necrotic
local lesions after 5-7 days (Fig. 3); no systemic
infection.
- C. quinoa. Small chlorotic local lesions after 4-6 days, subsequently becoming more diffuse; followed by systemic chlorotic leaf-spotting and apical stem necrosis (Fig. 1).
- Gomphrena globosa. Small grey necrotic local lesions after 5-6 days, later developing a red border and expanding to 4-5 mm (Fig. 2); no systemic infection.
- Hibiscus cannabinus (kenaf). Faint to fairly conspicuous systemic leaf chlorosis after 10-15 days; leaves produced subsequently are often symptomless.
- Nicotiana clevelandii. Faint systemic leaf chlorosis after 10-12 days.
- Trifolium incarnatum (crimson clover). Severe systemic leaf chlorosis after 15-18 days.
- C. quinoa. Small chlorotic local lesions after 4-6 days, subsequently becoming more diffuse; followed by systemic chlorotic leaf-spotting and apical stem necrosis (Fig. 1).
- Propagation species
- Nicotiana clevelandii and
Hibiscus cannabinus are useful for maintaining
cultures, and N. clevelandii is a
good source of virus for purification.
- Assay species
- Chenopodium murale is a sensitive and reliable local lesion assay host.
Strains
None reported.
Transmission by Vectors
Not transmitted by the aphid Myzus persicae (Brunt et al., 1980).
Transmission through Seed
Not seed-borne in Chenopodium quinoa or Nicotiana clevelandii (Brunt et al., 1980).
Serology
The virus is a good immunogen. Antiserum with a titre of 1/8192 in double-diffusion gel tests was obtained from a rabbit 33-53 days after a single intramuscular injection of c. 2 mg of virus emulsified with Freund’s complete adjuvant (Brunt et al., 1980).
Relationships
Although its properties indicate that hibiscus latent ringspot virus is a possible member of the nepovirus group, it is serologically unrelated to the following fifteen recognised or possible members of the group: arabis mosaic, arracacha A, artichoke Italian latent, cherry leaf roll, cherry rasp leaf, cocoa necrosis, grapevine Bulgarian latent, grapevine chrome mosaic, lucerne Australian latent, peach rosette mosaic, raspberry ringspot, strawberry latent ringspot, tomato black ring, tobacco ringspot and tomato ringspot viruses (Brunt et al., 1980). It is serologically unrelated also to 13 multi-component viruses of other groups, including okra mosaic virus (tymovirus group), which occurs naturally in H. rosa-sinensis in the Ivory Coast (Givord, 1974, 1977).
Stability in Sap
In Nicotiana clevelandii sap, the virus loses infectivity after 10 min at 64-66°C, 21-28 days at 20°C, 30-35 weeks at 2°C or dilution to 10-4 - 10-5.
Purification
Purified preparations of the virus are best obtained from Nicotiana clevelandii. Yields of up to 65 mg per kg leaf tissue have been obtained by the following procedure (Brunt et al., 1980). Extract systemically infected leaves (1 g/2 ml) in 0.07 M phosphate buffer at pH 7.6 containing 0.014 M mercaptoacetic acid, and clarify by adding n-butanol to 8.5% (v/v). Subject extracts to one cycle of differential centrifugation (20 min at 12,000 g; 90 min at 65,000 g), and resuspend the partially purified virus in 0.03 M neutral phosphate buffer (1 ml/40 g leaf material). Such preparations can be further purified by permeation chromatography on columns of controlled-pore glass beads (70 nm pores; 120-200 mesh) as described by Barton (1977).
The three components can be separated by centrifuging preparations on sucrose density gradient columns (10-40% w/v sucrose in 0.07 M phosphate buffer at pH 7.6) for up to 4.5 h at 85,000 g. However, complete separation of M (114 S) and B (132 S) component particles requires at least three cycles of density gradient and high speed centrifugation.
Properties of Particles
The particles, although of the same size, sediment as three components; apparently empty protein shells (T) and two nucleoproteins containing either 38% (M) or 42% (B) nucleic acid (Brunt et al., 1980).
Sedimentation coefficients (s20,w): at infinite dilution: 51 S (T), 114 S (M) and 132 S (B).
Buoyant density in CsCl (g/cm3): 1.32 (T), 1.49 (M) and 1.52 (B).
Estimated particle weights (x 10-6): 3.22 (T), 5.42 (M) and 5.82 (B).
A260/A280: 1.80 (unfractionated virus), 1.78 (M) and 1.85 (B).
A240/A260: 1.51 (unfractionated virus), 1.41 (M) and 1.56 (B).
Particle Structure
The particles are isometric, c. 28 nm in diameter and many are slightly angular in outline (Fig. 4, Fig. 5).
Particle Composition
Nucleic acid: Single-stranded RNA. The genome consists of two RNA molecules (RNA-1 and RNA-2) with M. Wt, estimated by polyacrylamide gel electrophoresis in non-denaturing conditions, of 2.5 x 106 and 2.2 x 106, respectively. B component particles contain one molecule of RNA-1 and M component particles one molecule of RNA-2.
Protein: The coat protein subunits have a M. Wt of 53,600 (estimated by electrophoresis in polyacrylamide gels containing sodium dodecyl sulphate).
Relations with Cells and Tissues
No information.
Notes
Although its vector is unknown, hibiscus latent ringspot virus has properties typical of nepoviruses. The sedimentation coefficient (114 S) and M. Wt of its M component particles indicate that it is especially similar to lucerne Australian latent, cherry leaf roll, grapevine Bulgarian latent, peach rosette mosaic and tomato ringspot viruses (Harrison & Murant, 1977; Jones, Forster & Mohamed, 1979) and might, therefore, be transmissible by nematodes (e.g. one or more species of Xiphinema).
Hibiscus latent ringspot virus is readily distinguished from two other viruses (hibiscus chlorotic ringspot and okra mosaic) known to infect Hibiscus rosa-sinensis (Waterworth, Lawson & Monroe, 1976; Waterworth, 1980; Givord, 1974, 1977). Although the three viruses induce similar symptoms in some malvaceous species and have isometric particles c. 28 nm in diameter, they are serologically distinct; moreover, the particles of hisbiscus latent ringspot virus sediment as three components (51, 115 and 132 S), whereas those of okra mosaic virus sediment as two components (42 and 106 S) and those of hibiscus chlorotic ringspot virus as a single component (118 S).
Figures

Leaf chlorosis and apical stem necrosis in systemically infected Chenopodium quinoa.
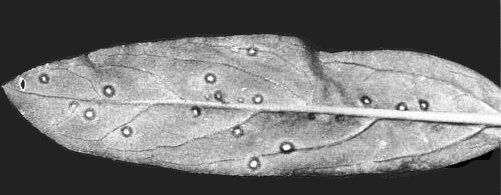
Necrotic lesions in inoculated leaf of Gomphrena globosa.

Necrotic lesions in inoculated leaf of Chenopodium murale.
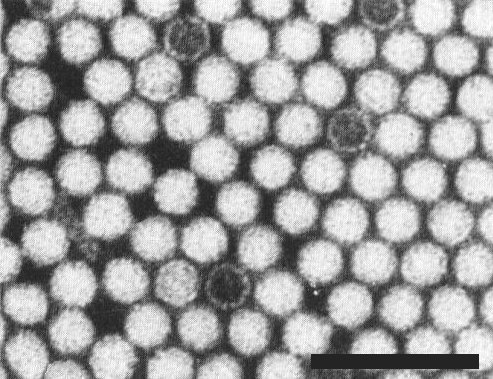
Virus particles in unfractionated preparation, Particles mounted in neutral potassium phosphotungstate. Bar represents 100 nm.

Virus particles in purified preparation of "bottom" component. Particles mounted in neutral potassium phosphotungstate. Bar represents 100 nm.
References list for DPV: Hibiscus latent ringspot virus (233)
- Barton, J. gen. Virol. 35: 77, 1977.
- Brunt, Barton, Phillips & Lana, Ann. appl. Biol. 96: 37, 1980.
- Givord, CMI/AAB Descriptions of Plant Viruses 128, 4 pp., 1974.
- Givord, Annls Phytopath. 9: 53, 1977.
- Harrison & Murant, CMI/AAB Descriptions of Plant Viruses 185, 4 pp., 1977.
- Jones, Forster & Mohamed, Ann. appl. Biol. 92: 49, 1979.
- Waterworth, CMI/AAB Descriptions of Plant Viruses 227, 4 pp., 1980.
- Waterworth, Lawson & Monroe, Phytopathology 66: 570, 1976.