Details of DPV and References
DPV NO: 234 July 1981
Family: Luteoviridae
Genus: Unassigned Luteoviridae
Species: Tobacco necrotic dwarf virus | Acronym: TNDV
Tobacco necrotic dwarf virus
S. Kubo Central Research Institute,The Japan Tobacco & Salt Public Corporation, Midori-ku, Yokohama, Japan
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
Described by Kubo, Tamura & Fukuda (1976) and Kubo & Takanami (1977).
A virus with RNA-containing isometric particles c. 25 nm in diameter occurring in phloem cells. Transmitted by aphids in the persistent manner, but not by inoculation with sap. It has a narrow host range. Recorded only from Japan.
Main Diseases
Causes stunting, and premature yellowing and death of the lower and intermediate-aged leaves, of tobacco (Nicotiana tabacum) plants (Fig. 1). Causes faint vein yellowing in spinach.
Geographical Distribution
Recorded only from Japan.
Host Range and Symptomatology
Twenty species in five families are known to be susceptible; solanaceous plants are the main hosts. Natural hosts are tobacco, spinach and Capsella bursa-pastoris. Spinach is the primary overwintering host. Gomphrena globosa and Claytonia perfoliata can be infected experimentally. Severity of phloem necrosis in tobacco plants varies between cultivars and is most prominent in cv. MC 1 (Fig. 1,Fig. 2).
- Diagnostic species
- Datura stramonium. Systemically infected leaves show
interveinal chlorosis.
- Nicotiana sylvestris. Systemically infected leaves show vein yellowing followed by interveinal chlorosis. Plants are severely stunted. Leaves bearing veinal necrosis roll downward and are malformed (Fig. 3).
- Physalis floridana. Interveinal chlorosis and moderate rolling of systemically infected leaves (Fig. 4).
- Nicotiana sylvestris. Systemically infected leaves show vein yellowing followed by interveinal chlorosis. Plants are severely stunted. Leaves bearing veinal necrosis roll downward and are malformed (Fig. 3).
- Propagation species
- Physalis floridana is suitable for maintaining cultures
and is the best source of virus for purification.
- Assay species
- Nicotiana sylvestris is useful for insect transmission tests. Symptoms appear about 10 days after infection (Fukuda, 1978).
Strains
Strains of the virus have not been studied in detail, but several isolates are distinguishable from the type strain on the basis of severity of reactions of Nicotiana spp. or Physalis floridana (S. Kubo, unpublished results).
Transmission by Vectors
The virus is transmitted by the aphid Myzus persicae in the persistent manner (Kubo et al., 1976). Experimentally, Acyrthosiphon solani also transmits the virus, but less efficiently (M. Tamura, unpublished results). M. persicae seems to be the only species that transmits the virus from the overwintering host (spinach) to tobacco plants. Minimum acquisition access period is 3 h and minimum inoculation access period is 30 min. There is a latent period in the vector of 2 to 4 days, depending on the dose of virus acquired. M. persicae injected with purified virus at high concentrations can transmit in 17 h. The aphid retains ability to transmit the virus after moulting and for periods longer than 30 days. In M. persicae injected with purified virus preparations, the virus particles are observed in the basal lamina of salivary glands including the accessory ones and in abdominal connective tissue cells, but there is no evidence for multiplication of the virus in the aphids (Kuwata & Kubo, 1978).
Transmission through Seed
Not seed-transmitted in N. tabacum.
Serology
Strongly immunogenic in rabbits. Antiserum with a titre of 1/4096 (microprecipitin test) has been obtained by intramuscular and intravenous injection (Kubo & Takanami, 1979). The conventional agar-gel double diffusion test is useful but, because of the low concentration of the virus in plants, concentrated or partially purified preparations must be used.
Relationships
Serologically, the virus is closely related to potato leafroll virus and distantly related to soybean dwarf and carrot red leaf viruses (Kubo & Takanami, 1978; Roberts, Tamada & Harrison, 1980; Waterhouse & Murant, 1981; S. Kubo, unpublished results). Tobacco necrotic dwarf and potato leafroll viruses both have a single-stranded RNA of M. Wt 2.0 x 106 and they share many properties with other members of the luteovirus group (Takanami & Kubo, 1979b).
Stability in Sap
Determined by inoculation of tobacco mesophyll protoplasts with partially purified virus preparations. Thermal inactivation point (10 min) is c. 80°C; infectivity is retained for more than 6 months at 4°C (Kubo & Takanami, 1980).
Purification
The following enzyme-assisted method is useful (Takanami & Kubo, 1979a). Powder frozen leaves of infected P. floridana, and disrupt in a mechanical blender in 0.1 M sodium citrate buffer (2 ml/g leaf), pH 6.0, containing 1% Driselase (an enzyme preparation which macerates plant tissue), and 0.1% thioglycollic acid. Shake the suspension for 2 h at 25 to 28°C and then emulsify by adding 0.5 vol. of a 1:1 (v/v) mixture of chloroform and n-butanol, and centrifuge at low speed. Precipitate the virus from the aqueous phase by adding polyethylene glycol (M. Wt 6000) to 8% (w/v) and NaCl to 0.4 M. Resuspend the virus in 0.01 M borate buffer, pH 8.0, concentrate by two cycles of differential centrifugation using 0.0l M phosphate buffer, pH 7.6 to 8.0, as the resuspending medium. Further purification is achieved by sucrose density gradient centrifugation. Virus yields are 5 to 10 mg per kg leaf material, at least ten times more than those obtained by methods that do not include the use of Driselase.
Properties of Particles
Purified virus preparations contain a single sedimenting component. The virus particles are disrupted in water and in the presence of low concentrations of sodium dodecyl sulphate or high concentrations of caesium salts (chloride, sulphate and formate). The particles precipitate reversibly when kept at 4°C in 0.01M phosphate buffers with pH values below 7.6 (Takanami & Kubo, 1979a, 1979b; Kubo & Takanami, 1980).
Sedimentation coefficient (s20,w): 115 S.
Isoelectric point: c. pH 5.3 (determined by electrofocusing).
Absorbance at 260 nm (1 mg/ml, 1 cm light path): 7.0 (not corrected for light-scattering).
A260/A280: 1.80 (not corrected for light-scattering).
Particle Structure
Particles are isometric, c. 25 nm in diameter in negatively-stained preparations (Fig. 6).
Particle Composition
Nucleic acid: Single-stranded RNA of M. Wt c. 2.0 x 106, estimated by electrophoresis in polyacrylamide gels in non-denaturing conditions. Two minor RNA components of M. Wt about 1 x 105 are also detected (Takanami & Kubo, 1979b).
Protein: The coat protein has a M. Wt of c. 25,700 daltons, estimated by electrophoresis in polyacrylamide/SDS gels (Kubo & Takanami, 1980). Each subunit of the coat protein contains c. 219 amino acid residues: ala, 12; arg, 21; asx, 20; cys, 3; glx, 16; gly, 23; his, 4; ile, 10; leu, 23; lys, 10; met, 2; phe, 9; pro, 9; ser, 24; thr, 12; trp, 2; tyr, 6; val, 13 (S. Kubo, unpublished results).
Relations with Cells and Tissues
The virus particles are confined to phloem tissue, namely sieve tubes, phloem parenchyma and companion cells (Kubo, Kuwata & Imaizumi, 1978; Imaizumi & Kubo, 1980a). They are found in nuclei, cytoplasm and vacuoles of infected cells (Fig. 5). Cytoplasm of the infected cells shows a fibrous appearance. Crystalline arrays of virus particles are not found. Despite its restriction to the phloem of intact plants, the virus can multiply in tobacco mesophyll protoplasts inoculated in vitro (Kubo & Takanami, 1979).
Epidermal cells of tobacco leaves have been infected with the virus by rubbing inoculation; the virus is thought not to spread from the primarily inoculated cells (Imaizumi & Kubo, 1980b).
Notes
The virus may be distinguished from potato leafroll, tobacco vein-distorting (Smith, 1946) and tobacco yellow-net viruses (Abeygunawardena, Karandawela & Bandaranayake, 1967) by the reactions of tobacco, P. floridana, and D. stramonium plants.
Figures

Disease outbreak caused by tobacco necrotic dwarf virus in tobacco cv. MC 1.

Severe phloem necrosis in MC 1 tobacco.

Stunting, interveinal chlorosis and malformation of leaves in Nicotiana sylvestris.

Physalis floridana: (left) healthy, (right) infected.
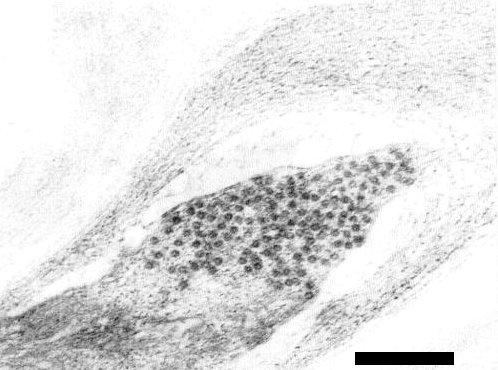
A group of virus particles in a section of a degenerate cell of Physalis floridana. Bar represents 200 nm.

Virus particles from a purified preparation stained with 2% phosphotungstate. Bar represents 100 nm.
References list for DPV: Tobacco necrotic dwarf virus (234)
- Abeygunawardena, Karandawela & Bandaranayake, Trop. Agriculturist 123: 37, 1967.
- Fukuda, Ann. Phytopath. Soc. Japan 44: 398, 1978.
- Imaizumi & Kubo, Ann. Phytopath. Soc. Japan 46: 54, 1980a.
- Imaizumi & Kubo, Ann. Phytopath. Soc. Japan 46: 422, 1980b.
- Kubo & Takanami, Ann. Phytopath. Soc. Japan 43: 76, 1977.
- Kubo & Takanami, Ann. Phytopath. Soc. Japan 44: 398, 1978.
- Kubo & Takanami, J. gen. Virol. 42: 387, 1979.
- Kubo & Takanami, Ann. Phytopath. Soc. Japan 46: 423, 1980.
- Kubo, Tamura & Fukuda, Hatabako Kenkyu 73: 49, 1976.
- Kubo, Kuwata & Imaizumi, Ann. Phytopath. Soc. Japan 44: 60, 1978.
- Kuwata & Kubo, Ann. Phytopath. Soc. Japan 44: 399, 1978.
- Roberts, Tamada & Harrison, J. gen. Virol. 47: 209, 1980.
- Smith, Parasitology 37: 131, 1946.
- Takanami & Kubo, J. gen. Virol. 44: 153, 1979a.
- Takanami & Kubo, J. gen. Virol. 44: 853, 1979b.
- Waterhouse & Murant, Ann. appl. Biol. 97: 191, 1981.