Details of DPV and References
DPV NO: 235 July 1981
Family: Virgaviridae
Genus: Pecluvirus
Species: Peanut clump virus | Acronym: PCV
Peanut clump virus
J.-C. Thouvenel Laboratoire de Virologie, Centre ORSTOM d'Adiopodoumé, B.P. V-51 Abidjan, Côte d’Ivoire
C. Fauquet Laboratoire de Virologie, Centre ORSTOM d'Adiopodoumé, B.P. V-51 Abidjan, Côte d’Ivoire
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
Described by
Thouvenel, Germani & Pfeiffer (1974) and
Thouvenel, Dollet & Fauquet (1976).
A virus with straight tubular particles of two lengths, containing single-stranded RNA. It is mechanically transmissible and has a relatively wide host range. It is seed-borne and soil-borne, and the fungus Polymyxa graminis is thought to be its vector. Found in West Africa.
Main Diseases
The common strain causes stunting (‘clump’) in groundnut (Arachis hypogaea); naturally infected groundnuts are patchily distributed in crops. A yellowing strain found in Upper-Volta (J. C. Thouvenel & C. Fauquet, unpublished results) causes concentric yellow line-patterns in old leaves but no stunting. Great millet (Sorghum arundinaceum) is a symptomless natural host (Dollet, Fauquet & Thouvenel, 1976).
Geographical Distribution
Found in West Africa (Ivory Coast, Senegal, Upper Volta).
Host Range and Symptomatology
Infects several species in the families Aizoaceae, Amaranthaceae, Chenopodiaceae, Cucurbitaceae, Gramineae, Leguminosae, Scrophulariaceae and Solanaceae. Mechanical transmission is difficult from Arachis hypogaea and Chenopodium amaranticolor because of inhibitors in the sap; purified preparations may be needed to obtain good transmission (Thouvenel & Fauquet, 1981). Typical symptoms are ringspots and line-patterns, and stunting of the plant.
-
Diagnostic species
- Arachis hypogaea
(groundnut). No local symptom, but mottle and chlorotic ringspots appear on new tip leaves 7-10 days after inoculation; these symptoms rapidly fade, then disappear when the leaves mature. The plant then stops growing and looks stunted (Fig. 1). With the yellow strain, mottle and chlorotic ringspots appear on new tip leaves and persist, the yellow colour becoming more prominent. -
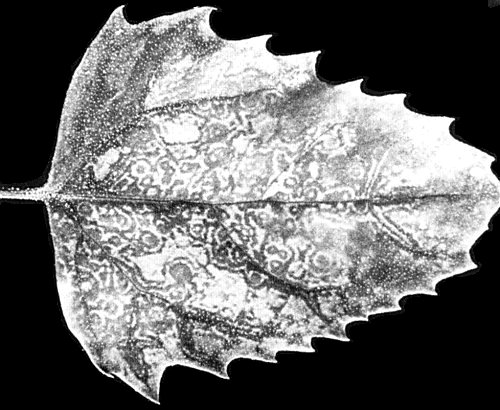
Chenopodium amaranticolor. Local yellow spots 2 days after inoculation, developing later
into ringspots and line-patterns which extend along the veins
(Fig. 2). Not systemic.
- Nicotiana benthamiana. Systemic mosaic and deformation (Fig. 3).
- Nicotiana glutinosa. Systemic mosaic.
- Phaseolus vulgaris (bean). Systemic mosaic.
- Triticum aestivum (wheat). Systemic mosaic and stunting (Fig. 4).
- Nicotiana benthamiana. Systemic mosaic and deformation (Fig. 3).
Propagation species
- Nicotiana benthamiana
and Phaseolus vulgaris are good sources of virus.Assay species
- Chenopodium amaranticolor
is a useful local lesion host; Sorghum arundinaceum is suitable for testing transmission to roots by vectors.
Strains
Most isolates of the virus from Senegal and Upper Volta differ little. An exceptional strain from Upper Volta causes yellowing symptoms in groundnut, completely different from the stunting induced by common strains, but its other properties are similar to those of the common strain and they are serologically indistinguishable.
Transmission by Vectors
The virus is thought to be transmitted by a plasmodiophoromycete fungus to graminaceous hosts and to groundnut. The infectivity of soil was related to the presence of Polymyxa graminis, resting spores of which were detected readily in roots of Sorghum arundinaceum bait plants that became infected with the virus (Fig. 5) (Thouvenel & Fauquet, 1980, 1981).
Transmission through Seed
Seed transmission is known only in groundnut (5-6%). Seeds collected from the diseased plants thus obtained (second generation seeds) produced c. 20% of diseased seedlings (Thouvenel, Fauquet & Lamy, 1978; Thouvenel & Fauquet, 1981).
Serology
Highly immunogenic. Antiserum with a titre of 1/2048 in microprecipitin tests can be obtained.
Relationships
Peanut clump virus resembles other rod-shaped viruses with known or suspected fungus vectors, such as beet necrotic yellow vein, potato mop-top and wheat soil-borne mosaic viruses, but did not react with antisera to these viruses or to tobacco mosaic, tobacco rattle, pea early-browning or nicotiana velutina mosaic viruses (Thouvenel et al., 1976; Thouvenel & Fauquet, 1981); antiserum to peanut clump virus did not react with barley stripe mosaic or tobacco mosaic viruses (Thouvenel & Fauquet, 1981). D. V. R. Reddy (personal communication) found no reaction between a virus associated with a ‘clump’ disease of groundnut in India and antiserum to peanut clump virus.
Stability in Sap
In C. amaranticolor sap the thermal inactivation point (10 min) is 64°C, and the dilution endpoint 10-4. Infectivity is retained for 4 weeks at 27°C, and for more than a year in frozen sap or frozen leaves (Thouvenel et al., 1976).
Purification
Grind frozen leaves in 0.5 M borate buffer (pH 8) containing reducing agents, and clarify the extract with butanol/chloroform (Thouvenel et al., 1976), or by acidification to pH 4.2 (Thouvenel, Fauquet & Dollet, 1978). Concentrate the virus particles by 2 or 3 cycles of differential centrifugation, resuspending the high speed pellets in 0.05 M borate buffer (pH 8). Additional purification can be obtained by isopycnic centrifugation in 45% CsCl. About 20-25 mg of virus may be obtained from 1 kg leaves.
Properties of Particles
The particles sediment as two components, with sedimentation coefficients (s20,w) of 183 and 224 S. There is no information about the infectivity of the two types of particle.
Isoelectric point: pH 6.45 ± 0.10.
Amin: 249 nm; Amax: 270 nm (corrected for light-scattering).
A260/A280 = 1.00; A270/A249 = 1.32.
Buoyant density in CsCl is about 1.32 g/cm3 (Thouvenel & Fauquet, 1981).
Particle Structure
Particles are straight rods, helically constructed with a hollow core. They are about 21 nm wide and of two predominant lengths: 190 and 245 nm (Fig. 6) (Thouvenel et al., 1976). The length distribution of the short particles is broad, and in some preparations an additional class of 160 nm particles is recognizable.
Particle Composition
Nucleic acid: RNA, single-stranded (determined by nuclease digestion), comprising about 4% of the weight of the particles (estimated from the absorption spectrum). In most RNA preparations, polyacrylamide gel electrophoresis reveals two components with estimated M. Wt of about 1.7 x 106 and 2.1 x 106. In a few preparations, the smaller component can be resolved into 2 or 3 components (M. Wt 1.65-1.75 x 106) (J. C. Thouvenel & C. Fauquet, unpublished results).
Protein: Electrophoresis of coat protein preparations in polyacrylamide gels reveals one polypeptide of M. Wt c. 23,000. Subunits have the following amino acid composition: asp 27 residues; thr 8; ser 13; glu 25; pro 12; gly 26; ala 21; cys ?; val 20; met 0; ile 11; leu 18; tyr 5; phe 7; his 5; lys 7; arg 19; trp ?; total 224 residues + cys + trp (J. C. Thouvenel & C. Fauquet, unpublished results).
Relations with Cells and Tissues
The virus is found in the cells of roots, stems and leaves of systemically infected graminaceous plants. Virus particles in wheat cells are found in the cytoplasm, near the nucleus or along the plasmalemma, and arranged in angled-layer aggregates (M. Dollet, personal communication).
Notes
The closest affinities of peanut clump virus are with beet necrotic yellow vein, broad bean necrosis, potato mop-top and soil-borne wheat mosaic viruses; it differs from all four in infecting Phaseolus vulgaris systemically. It is distinguishable from all other viruses of groundnut by its particle morphology, its antigenic specificity, its characteristic symptoms in C. amaranticolor and its wide host range (when inoculated in the form of purified virus). Clump disease is controlled by treating the soil with the fumigant nematicides dibromochlorophenol or DD, which may also have a fungicidal action (Germani, Thouvenel & Dhéry, 1975). It is not controlled by the use of systemic nematicides.
Figures

Symptoms of the clump disease of groundnut in the field.
Symptoms in an inoculated leaf of Chenopodium amaranticolor.

Systemic symptoms in Nicotiana benthamiana.

Systemic symptoms in Triticum aestivum infected by mechanical inoculation (left); healthy plant (right).
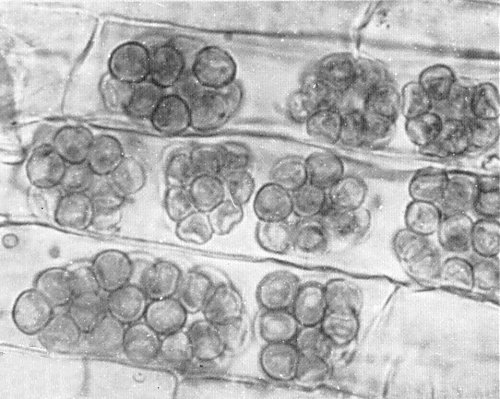
Resting spores of Polymyxa graminis in Sorghum arundinaceum roots.

Electron micrograph of purified virus particles, stained with uranyl acetate. Bar represents 200 nm.
References list for DPV: Peanut clump virus (235)
- Dollet, Fauquet & Thouvenel, Pl. Dis. Reptr 60: 1076, 1976.
- Germani, Thouvenel & Dhéry, Oléagineux 30: 259, 1975.
- Thouvenel & Fauquet, Pl. Dis. 64: 957, 1980.
- Thouvenel & Fauquet, Ann. appl. Biol. 97: 99, 1981.
- Thouvenel, Germani & Pfeiffer, C. r. hebd. Séanc. Acad. Sci., Paris 278: 2847, 1974.
- Thouvenel, Dollet & Fauquet, Ann. appl. Biol. 84: 311, 1976.
- Thouvenel, Fauquet & Dollet, Annls Phytopath. 10: 93, 1978.
- Thouvenel, Fauquet & Lamy, Oléagineux 33: 503, 1978.