Details of DPV and References
DPV NO: 236 July 1981
Family: Bromoviridae
Genus: Bromovirus
Species: Melandrium yellow fleck virus | Acronym: MYFV
Melandrium yellow fleck virus
M. Hollings Glasshouse Crops Research Institute, Littlehampton, Sussex, BN16 3PU, England
J. Horváth University of Agricultural Sciences, Institute for Plant Protection, H-8361 Keszthely, P.O. Box 71, Hungary
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
Described by Hollings & Horváth (1978), and Hollings, Horváth & Besada (1978).
A virus with RNA-containing isometric particles c. 25 nm diameter, found in Melandrium album in Hungary. The virus is readily sap-transmissible to a wide range of herbaceous plant species. No vector is known.
Main Diseases
In naturally infected or inoculated Melandrium album, the virus induces systemic vein clearing and yellow flecks. Experimental infection causes mosaic diseases in beet (Beta spp.) and cucumber (Cucumis sativus), and lethal necrosis in pea (Pisum sativum).
Geographical Distribution
Known only from Hungary.
Host Range and Symptomatology
The virus infected 173 of 179 species in 17 of 18 plant families (not Cruciferae). The virus systemically infected 71 of 73 species in the Caryophyllaceae, but it induced only local lesions in Dianthus barbatus and did not infect virus-free carnation (D. caryophyllus) cv. William Sim.
- Diagnostic species
No species gives diagnostic reactions, but a combination of the following species is useful:
Chenopodium quinoa. Numerous small, necrotic brown local lesions in 3-5 days (Fig. 2), enlarging and coalescing to give necrotic areas. No systemic infection.
Cucumis sativus cv. Butcher’s Disease Resister. Sharply-defined yellowish local lesions in 4-7 days; occasional systemic infection, with chlorotic spots and flecks.
Melandrium album. Local necrotic rings after 5-7 days; systemic vein clearing and yellow flecks.
Nicotiana clevelandii. Chlorotic and necrotic local lesions in 3-5 days, becoming extensive necrotic areas. Systemic severe mottle, blistering, puckering, rosetting and much necrosis (Fig. 1).
Phaseolus vulgaris (French bean) cv. The Prince. Numerous small brown necrotic local lesions in 3 days (Fig. 3); no systemic infection.
Reactions in other commonly used test plants include:
Chenopodium amaranticolor, C. capitatum, C. murale. Necrotic local lesions (Fig. 4). Systemic infection occasionally occurs in C. murale, with leaf dwarfing and chlorosis.
Datura stramonium. Local chlorotic spots and rings in about 1 week; no systemic infection.
Gomphrena globosa. Small, whitish necrotic local lesions in about 1 week; systemic coarse yellowish mottle and stunting.
Lycopersicon esculentum (tomato) cv. Moneymaker. Few chlorotic local lesions in about 1 week; no systemic infection.
Nicotiana glutinosa. Few chlorotic local lesions; no systemic infection.
N. glutinosa x N. clevelandii hybrid. Chlorotic local lesions after about 1 week; systemic bright yellow-green mottle in 2 weeks, with some dwarfing. Subsequent growth with only slight mottle.
N. tabacum (tobacco) White Burley. Chlorotic local lesions in 7-10 days, enlarging to chlorotic rings and ring-spots. No systemic infection.
Petunia hybrida cv. Fire Chief. A few chlorotic local lesions in 5-6 days; no systemic infection.
Pisum sativum (pea) cv. Onward. Systemic severe mottle, leaf distortion and stunting, with spreading necrosis killing the plant.
Tetragonia expansa. White necrotic local lesions after about 6 days; systemic vein yellowing, leaf twisting and malformation.
Tropaeolum majus. Irregular chlorotic local lesions and areas; systemic yellow vein-netting and leaf dwarfing.
- Propagation species
N. clevelandii is useful for virus propagation and for maintaining cultures.
- Assay species
Chenopodium quinoa and Phaseolus vulgaris give satisfactory local lesions.
Strains
No information.
Transmission by Vectors
No transmission was obtained by the aphids Myzus persicae or Macrosiphum euphorbiae, given either long or short feeds (Hollings & Horváth, 1978, and unpublished information).
Transmission through Seed
No seed transmission was detected in N. clevelandii.
Serology
The virus is a rather poor immunogen, but a good antigen in vitro. Antisera prepared in rabbits by one intravenous injection, followed after 1 and 3 weeks by two intramuscular injections (with Freund’s complete adjuvant), with about 1 mg virus/injection, had specific titres of 1/512 in tube precipitin tests. Somatic (granular) precipitates were produced.
In immunodiffusion, a single line of precipitation is formed (Fig. 5), and good reactions were obtained in 0.8% agar or agarose gels in distilled water or in 0.02 M phosphate buffer (Hollings & Horváth, 1978).
Relationships
The virus has properties in vitro typical of bromoviruses, but no serological relationship was detected in reciprocal tests with brome mosaic, broad bean mottle or cowpea chlorotic mottle viruses (M. Hollings, unpublished information). Nor was any serological reaction detected, in reciprocal immunodiffusion tests, to 70 strains of 50 RNA-containing viruses of similar morphology, representing at least 12 taxonomic groups (Hollings & Horváth, 1978, and unpublished information).
Stability in Sap
The virus is very stable in sap of Nicotiana clevelandii. It was still infective at dilutions of 10-5 and withstood 10 min at 85° but not 90°C. Infectivity survived 3 but not 5 weeks at 20°C and at least 1 year at 2°C. Lyophilised sap containing 7% (w/v) each of dextrose and peptone was highly infective after 4 years under vacuum at about 20°C (M. Hollings, unpublished information), and purified preparations survived at least 3 years at 2°C. Infectivity was only slightly diminished by treatment with diethyl ether (20% v/v, 24 h, 4°C), and the virus was stable in Tris buffer at pH 8.5 in the presence of 0.001 M EDTA and 1 M NaCl (Hollings & Horváth, 1978).
Purification
The virus is readily purified from systemically infected N. clevelandii (Hollings & Horváth, 1978): harvest plants 10-14 days after infection, homogenise in 0.02 M phosphate buffer, pH 7.6, (4 ml/g leaf) containing 0.1% v/v thioglycollic acid. Filter through cloth, add n-butanol (9.3 ml/100 ml juice) and stir for 1 h at 20°C. Concentrate the virus by one or more cycles of differential centrifugation (20 min at 12,000 g : 90 min at 80,000 g). Re-suspend the final pellets in 0.03 M phosphate buffer (1 ml/15 g original leaf) and, after about 3 h, remove insoluble material by brief centrifugation (10 min, 10,000 g).
Further purification can be achieved by molecular permeation chromatography: apply 1.0-1.5 ml of the partially purified preparations to columns (85 x 1.5 cm) of controlled-pore glass beads (70 nm pore size), and elute the virus with 0.05 M Na-phosphate buffer, pH 7.6. The virus is eluted immediately after the void volume (Barton, 1977; M. Hollings & R. J. Barton, unpublished results). Yields of 200 mg virus/kg leaf can be obtained.
Properties of Particles
Purified preparations produce one light-scattering band in sucrose density-gradient centrifugation; infectivity, serological activity and virus particles are almost confined to this band. In analytical ultracentrifugation, one major sedimenting component (s20, w = 88 S) and sometimes one minor component (56 S) were present (M. Hollings, unpublished information).
A260/A280: 1.66 to 1.69; Amax(260)/Amin(242): 1.39 to 1.43 (values corrected for light-scattering).
Electrophoretic behaviour: in immunoelectrophoresis in 0.8% Ionagar no. 2 in 0.02 M phosphate buffer pH 7.6 at 2°C, a single antigenic component was present, moving rather rapidly towards the anode (Fig. 5) at approx. 9.5 x 10-5 cm2 v-1 sec-1.
Particle Structure
In 2% phosphotungstate pH 6.5, particles are isometric, about 25 nm diameter (Fig. 6) (Hollings & Horváth, 1978). Some purified preparations also contain small isometric particles about 17 nm diameter, and tubular structures of about the same diameter as the virus particles and of irregular length (Fig. 6) (M. Hollings & P. T. Atkey, unpublished information). Both these structures resemble those recorded for cowpea chlorotic mottle virus (Bancroft et al., 1967).
Particle Composition
Nucleic acid: RNA, single-stranded, of M. Wt 1.2, 1.1, 1.0 and 0.3 x 106, comprising about 20% of particle weight (estimated spectrophotometrically) (Barton & Hollings, 1981).
Protein: Polyacrylamide gel electrophoresis showed a single polypeptide of M. Wt 22,000 ± 600 (Barton & Hollings, 1981), contrasted with 20,300 + 800 for brome mosaic virus.
Relations with Cells and Tissues
The virus is present in leaves, stems and roots of infected Nicotiana clevelandii and Melandrium album.
Notes
The virus is readily identified by transmission to N. clevelandii and immunodiffusion tests with crude N. clevelandii sap. Serologically specific electron microscopy greatly enhances the sensitivity of detection. MYFV appears to be a distinct member of the bromovirus group.
Figures

Nicotiana clevelandii plants: (left) 7 days after infection, showing irregular necrotic local lesions, and developing systemic leaf mottle and puckering; (right) 20 days after infection, showing very severe stunting, rosetting, puckering, mottle and leaf necrosis.

Chenopodium quinoa: necrotic local lesions 5 days after infection.
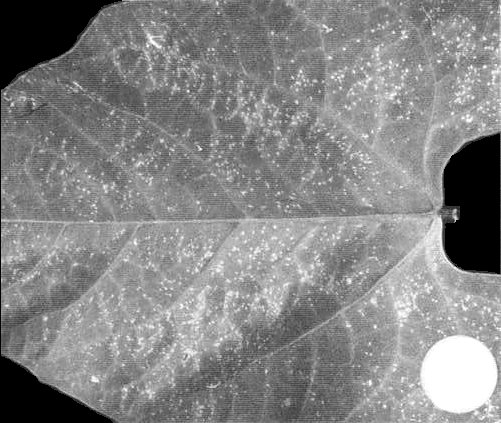
Phaseolus vulgaris cv. The Prince: necrotic dot local lesions 11 days after infection.

Chenopodium murale: necrotic local lesions 8 days after infection.
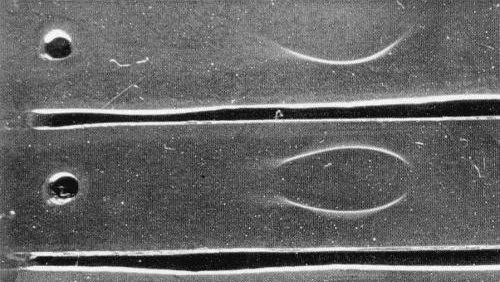
Immunoelectrophoretogram: after 1 h electrophoresis in 0.8% (w/v) Ionagar no. 2 in 0.02 M phosphate buffer (pH 7.6) at 2°C, 4.4 v/cm. The single antigenic component migrates rather rapidly towards the anode.
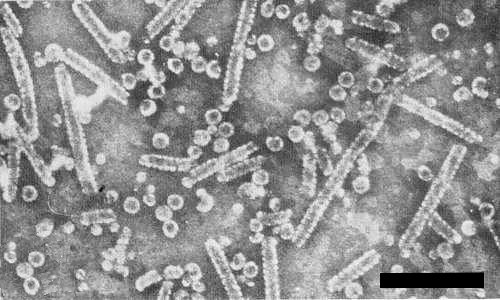
Partially purified preparation of the virus stained in phosphotungstate (pH 6.5), showing isometric particles and tubular structures. Bar represents 200 nm.
References list for DPV: Melandrium yellow fleck virus (236)
- Bancroft, Hills & Markham, Virology 31: 354, 1967.
- Barton, J. gen. Virol. 35: 77, 1977.
- Barton & Hollings, Abstr. 5th int. Congr. Virology, Strasbourg, 1981: W22, 1981.
- Hollings & Horváth, Rep. Glasshouse Crops Res. Inst. for 1977: 129, 1978.
- Hollings, Horváth & Besada, Abstr. 3rd int. Congr. Plant Path., Munich, 1978: 26, 1978.