Details of DPV and References
DPV NO: 238 July 1981
Family: Secoviridae
Genus: Comovirus
Species: Quail pea mosaic virus | Acronym: QPMV
Quail pea mosaic virus
B. J. Moore Department of Plant Pathology, University of Arkansas, Fayetteville, AR 72701, USA
H. A. Scott Department of Plant Pathology, University of Arkansas, Fayetteville, AR 72701, USA
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
Described by
Moore (1973).
Synonym
- Bean curly dwarf mosaic virus (Rev. Pl. Path. 56: 5373) is regarded as a strain
-
A virus with isometric particles about 30 nm in diameter, sedimenting as three components. It is transmitted by beetles and is easily sap transmissible. Occurs in USA (Arkansas) and Central America.
Main Diseases
Causes mosaic in quail pea (Strophostyles helvola; Fig. 1), soybean (Glycine max; Fig. 3) and bean (Phaseolus vulgaris).
Geographical Distribution
Found in USA (Arkansas) and Central America. Occurs naturally in quail pea and soybean in Arkansas and in Phaseolus bean in El Salvador and Costa Rica.
Host Range and Symptomatology
Transmitted to 23 species in three plant families, including 21 species in the Leguminosae and one each in the Chenopodiaceae and Convolvulaceae.
-
Diagnostic species
- Ipomoea nil
(morning glory) cv. Scarlet O’Hara. Necrotic local lesions. No systemic infection. - Phaseolus vulgaris (French bean) cvs Pinto and Topcrop. Diffuse
necrotic local lesions
(Fig. 2).
The type (quail pea mosaic) strain causes no
systemic infection but the bean curly dwarf mosaic strain infects both cultivars
systemically.
- Vigna unguiculata (cowpea) cv. Monarch. The type strain causes chlorotic local lesions with no systemic infection.
- Gomphrena globosa (globe amaranth). Infected systemically by the bean curly dwarf mosaic strain. No infection by the type strain.
- Vigna unguiculata (cowpea) cv. Monarch. The type strain causes chlorotic local lesions with no systemic infection.
-
Propagation species
- Phaseolus vulgaris
cvs Black Valentine and Cherokee Wax, and Glycine max (soybean) cv. Kanrich are systemically infected with both strains and are good sources of virus for purification.Assay species
- Phaseolus vulgaris
cv. Pinto is satisfactory for local lesion assays of the type strain. Dolichos lablab is used for the bean curly dwarf mosaic strain. - Glycine max cv. Lee has been used for studies on vector transmission.
Strains
The type (quail pea mosaic) strain was collected in Arkansas, USA (Moore, 1973). Isolates from soybean in Arkansas and from Phaseolus bean in Costa Rica have not been compared with the type strain in host range and symptomatology but are serologically distinguishable from it (Fulton & Scott, 1979). Bean curly dwarf mosaic virus, collected in El Salvador (Meiners et al., 1977), differs from the type strain in host range and symptomatology, but is serologically closely related to it and is regarded as a strain.
Transmission by Vectors
The type strain is transmitted by the bean leaf beetle, Ceratoma trifurcata (Chrysomelidae) (Moore, 1973). The bean curly dwarf mosaic strain is transmitted by the spotted cucumber beetle, Diabrotica undecimpunctata (Chrysomelidae), and the Mexican bean beetle, Epilachna varivestis (Coccinellidae) (Meiners et al., 1977).
Transmission through Seed
The type strain was not transmitted through seed of Black Valentine or Cherokee Wax bean (Moore, 1973). No seed transmission of the bean curly dwarf mosaic strain was detected in nine cultivars of Phaseolus vulgaris or one cultivar of P. acutifolius (Meiners et al., 1977).
Serology
The virus is strongly immunogenic. Antiserum titres of 1/1024 are easily obtained (Moore, 1973). Antiserum with a titre of 1/128 was obtained against the bean curly dwarf mosaic strain (Meiners et al., 1977). Good reactions are observed in gel-diffusion tests with crude sap.
Relationships
Quail pea mosaic virus has particle properties typical of comoviruses and is related serologically to other members of the comovirus group. It is the type member of one of the five comovirus serogroups proposed by Fulton & Scott (1979) and is distantly related to viruses in each of the other four serogroups. Fig. 6 shows distant relationships with the type members of three of the serogroups: bean pod mottle virus, bean rugose mosaic virus and cowpea severe mosaic virus.
Stability in Sap
In soybean sap, particles of the type strain lose infectivity after heating for 10 min at 55° to 60°C, dilution between 10-3 and 10-4, and after incubation for between 5 and 6 days at 22°C (Moore, 1973). Infectivity of particles of the bean curly dwarf mosaic strain in bean sap is abolished by heating for 10 min between 50° and 55°C, dilution between 10-5 and 10-6, and after incubation for between 3 and 4 weeks at 22°C (Meiners et al., 1977).
Purification
1. Type strain
(Moore, 1973).
Harvest infected plants 10-18 days after
inoculation, freeze, then homogenize with 0.2 M phosphate buffer, pH 7.2
(2 ml per g tissue). Express sap through cheesecloth, adjust to pH 5.0 with
0.1 M HCl, stir at room temperature for 30 min, centrifuge at low speed and
subject the supernatant fluid to alternate cycles of high- and low-speed
centrifugation. Resuspend the virus particles in 0.01 M phosphate buffer,
pH 7.2.
2. Type strain (B. J. Moore & H. A. Scott, unpublished data). Virus
particles can be purified by a modification of Steere’s chloroform-butanol
method. Extract each 1 g tissue in 2 ml buffer (0.2 M phosphate + 0.1 M
ascorbic acid, pH 7.2) and 2 ml of a 1:1 mixture of chloroform and butanol.
Subject the aqueous phase to 3 cycles of alternate high and low-speed
centrifugation. Resuspend the virus particles in 0.01 M phosphate buffer,
pH 7.2.
3. Bean curly dwarf mosaic strain (Meiners et al., 1977). Harvest infected plants 14 days after inoculation. Extract each 1 g fresh bean leaves in 2 to 3 ml buffer (0.02 M sodium citrate buffer, pH 7.5 + 0.02 M 2-mercaptoethanol) and 0.2 ml cold chloroform. Centrifuge at low speed for 10 min. Concentrate the virus from the supernatant fluid by high speed centrifugation or by precipitation with 10% (w/v) polyethylene glycol, M. Wt 6000. Resuspend the virus pellets in 0.02 M citrate buffer, pH 7.5. Coloured impurities can be removed by adding activated charcoal at 0.05 g/ml and centrifuging at low speed. The virus in the supernatant fluid can be further purified by density gradient centrifugation.
Properties of Particles
Purified preparations of the virus contain three centrifugal components (Fig. 5).
Sedimentation coefficients (s20,w): 57, 93 and 112 S respectively for top, middle and bottom components (Moore, 1973).
Electrophoresis: unfractionated virus particles migrate as two immunoelectrophoretic components (Fig. 4) as reported for other comoviruses (Bruening, 1978).
A260/A280: 1.6 for unfractionated virus, uncorrected for light-scattering.
Particle Structure
Particles are isometric, c. 30 nm in diameter (Fig. 7). Some particles are penetrated by stain.
Particle Composition
Nucleic acid: Probably single-stranded RNA. Two nucleic acid species extracted from unfractionated purified preparations of virus particles co-migrated with RNA species from cowpea severe mosaic virus (De Jager, 1979) when subjected to polyacrylamide gel electrophoresis under non-denaturing conditions (H. A. Scott & B. J. Moore, unpublished data).
Protein: Polyacrylamide gel electrophoresis yields two proteins with M. Wt of c. 22,000 and 40,000 (H. A. Scott & B. J. Moore, unpublished data).
Relations with Cells and Tissues
No information.
Notes
Quail pea mosaic virus, a member of the comovirus group, may be distinguished from other legume viruses outside the comovirus group by its 30 nm isometric particles, its possession of three centrifugal components, and its transmission by beetles. It may be separated from other members of the comovirus group by its antigenic specificity (Fulton & Scott, 1979). The bean curly dwarf mosaic strain is reported to be widely distributed in beans in El Salvador (Meiners et al., 1977). The prevalence of the type strain in the USA is not known.
Figures

Mosaic in trifoliolate leaf of Strophostyles helvola.

Local lesions in primary leaf of Phaseolus vulgaris cv. Pinto.

Mosaic in trifoliolate leaf of Glycine max cv. Lee.
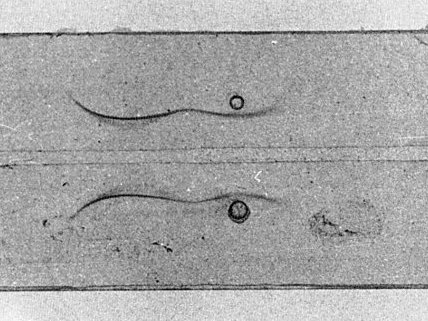
Immunoelectrophoresis of purified virus particles. Anode to the left.
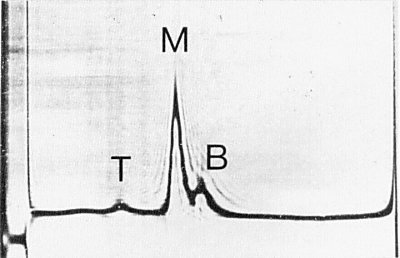
Schlieren diagram obtained by analytical centrifugation of a purified virus preparation showing top (T), middle (M) and bottom (B) components. Sedimentation is from left to right.

Immunodiffusion test comparing quail pea mosaic virus (QP) with bean pod mottle (HP), bean curly dwarf mosaic (BC), bean rugose mosaic (BR), and cowpea severe mosaic (CS) viruses. Central well contains antiserum to quail pea mosaic virus.

Virus particles stained with sodium phosphotungstate, pH 6.8. Bar represents 100 nm.
References list for DPV: Quail pea mosaic virus (238)
- Bruening, CMI/AAB Descriptions of Plant Viruses 199, 5 pp., 1978.
- De Jager, CMI/AAB Descriptions of Plant Viruses 209, 5 pp., 1979.
- Fulton & Scott, Phytopathology 69: 305, 1979.
- Meiners, Waterworth, Lawson & Smith, Phytopathology 67: 163, 1977.
- Moore, Pl. Dis. Reptr 57: 311, 1973.