Details of DPV and References
DPV NO: 240 July 1981
Family: Potyviridae
Genus: Potyvirus
Species: Leek yellow stripe virus | Acronym: LYSV
Leek yellow stripe virus
L. Bos Research Institute for Plant Protection, Wageningen, The Netherlands
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
- Disease first reported by Bremer (1937) and described by Kupke (1957), and virus identified and
named by Bos et al. (1978a).
- A virus with flexuous filamentous particles c. 820 nm long. Transmitted by aphid species in the non-persistent manner and by inoculation with sap. Infects leek (Allium porrum) and a few other Allium species.
- Widespread in several European countries, probably world-wide.
- A virus with flexuous filamentous particles c. 820 nm long. Transmitted by aphid species in the non-persistent manner and by inoculation with sap. Infects leek (Allium porrum) and a few other Allium species.
Main Diseases
In nature, the virus seems largely limited to leek (Allium porrum) in which it causes irregular yellow striping of the whole lamina, but especially at its base (Fig. 1, Fig. 2). Whole leaves may be yellow. Affected plants are less juicy and weigh less than normal, the leaves are slightly flaccid, the white stems are lustreless and the keeping quality of the harvested product is impaired. Infected plants in autumn and winter crops suffer more than healthy plants from frost and may be killed (Bos et al., 1978a). Leaf streaking is more white and severe when the plants are also infected with shallot latent virus (Paludan, 1980). All cultivars tested were sensitive (Bos et al., 1978a; Paludan, 1980).
Geographical Distribution
Reported from several European countries, most probably restricted to areas with year-round cultivation of leek.
Host Range and Symptomatology
Only 9 out of 32 Allium species tested were found susceptible and most were infected symptomlessly (Graichen, 1978). Onion (A. cepa) and shallot (A. ascalonicum) are very resistant but not immune (Bos et al., 1978a) and the virus has been isolated from plants of these species grown near severely infected leek (Graichen, 1978). A. fistulosum is immune (Bos et al., 1978a; Graichen, 1978). The virus produces local lesions in Chenopodium album, C. amaranticolor and C. quinoa (Bos et al., 1978a; Graichen, 1978) and in four other Chenopodium spp. and in Celosia argentea (Graichen, 1978) but does not infect 28 other dicotyledonous species (Verhoyen & Horvat, 1973; Bos et al., 1978a; Graichen, 1978).
- Diagnostic species
- Allium porrum
(leek). Distinct yellow or chlorotic striping of systemically infected leaves starting c. 14 days after inoculation. - Allium cepa (onion). Infected only with difficulty and then almost symptomless (this
distinguishes the virus from onion yellow dwarf virus).
- Chenopodium amaranticolor. Small chlorotic local lesions appear c. 3 weeks after inoculation and develop into green rings when the leaves turn yellow (Fig. 3). (Note that onion yellow dwarf virus does not infect this species and that lesions produced by shallot latent virus develop sooner, and are smaller and more necrotic). No systemic infection.
- Chenopodium quinoa. Chlorotic local lesions may appear c. 11 days after inoculation but more often become evident as green rings only when the leaves turn yellow (Fig. 4). No systemic infection.
- Chenopodium amaranticolor. Small chlorotic local lesions appear c. 3 weeks after inoculation and develop into green rings when the leaves turn yellow (Fig. 3). (Note that onion yellow dwarf virus does not infect this species and that lesions produced by shallot latent virus develop sooner, and are smaller and more necrotic). No systemic infection.
- Propagation species
- Allium porrum
is suitable for maintaining the virus and is a good source of virus for purification.- Assay species
- Chenopodium amaranticolor
and especially C. quinoa are moderately useful for local lesion assays.
Strains
None reported.
Transmission by Vectors
Transmitted experimentally by Aphis fabae (Verhoyen & Horvat, 1973) and Myzus persicae (Bos et al., 1978a) in the non-persistent manner.
Transmission through Seed
Not detected in 1000 seedlings each of 15 leek cultivars or in 24,000 seedlings of leek cv. Goliath grown from seed from virus-infected plants; in contrast, plantlets from inflorescence bulbils from such plants were all infected (Bos et al., 1978a).
Serology
D. Z. Maat prepared a specific antiserum with a titre in micro-precipitin tests of 1/4096 with purified virus, and of 1/256 with virus in clarified plant extracts (Bos et al., 1978a).
Relationships
The virus belongs to the potyvirus group. It is serologically related to, but distinguishable from onion yellow dwarf virus. An antiserum to leek yellow stripe virus had a titre of 1/4096 to purified homologous virus and 1/4 to onion yellow dwarf virus. An antiserum to onion yellow dwarf virus had a homologous titre of 1/1024 and a heterologous titre of 1/16 to leek yellow stripe virus (Bos et al., 1978a).
Stability in Sap
In leek sap, the thermal inactivation point is between 50 and 60°C. The dilution end-point is 10-2 to 10-3 and longevity in vitro is 3 to 4 days (Bos et al., 1978a; Graichen, 1978). The virus withstands drying and storage in leaves over CaCl2 at 4°C for at least 9 years (L. Bos, unpublished data).
Purification
Purification methods successful for other potyviruses, such as clarification with butanol/chloroform mixtures, butanol alone or Triton X-100. or precipitation with ammonium sulphate, failed (Verhoyen & Horvat, 1973) and the mucilage in sap of Allium spp. poses special problems (Huttinga, 1975). The following method (Huttinga, 1975; Bos et al., 1978a) proved useful:
Grind 100 g tissue in 500 ml 0.1 M Tris buffer (adjusted to pH 9 with thioglycollic acid) together with 20 ml chloroform, 20 ml carbon tetrachloride and 10 ml diethyl ether. Centrifuge for 10 min at 4000 g. Centrifuge the supernatant fluid for 1.5 h at 26,500 g and resuspend pellets in 50 ml 0.1 M Tris-HCl buffer pH 9. After 2 h at 4°C centrifuge for 10 min at 8000 g; then pass the filtrate through a Sephadex G-200 column with the above buffer containing 4 x 10-4 M NaN3, using a peristaltic pump to give a flow rate of 4.6 ml/h/cm2. Pool the virus-containing ultraviolet-absorbing fractions and concentrate by centrifuging for 1.75 h at 47,000 g. Resuspend the pellets, and centrifuge through a 10-40% sucrose density gradient for 2 h at 25,000 rev./min.
Properties of Particles
The particles sediment as a single infective component. Buoyant density of virus particles in CsCl: 1.326 g/cm3.
Particle Structure
The particles are flexuous, with a modal length of c. 815 (Verhoyen & Horvat, 1973) to 820 nm (Bos et al., 1978a). They usually aggregate end-to-end and cluster in crude sap (Fig. 6) (Verhoyen & Horvat, 1973; Bos et al., 1978a).
Particle Composition
M. Wt of coat protein subunit is c. 34,000 (Huttinga, 1975).
Relations with Cells and Tissues
Most epidermal cells contain one or two fibrous (sometimes granular) cytoplasmic inclusion bodies, often near the nucleus. They stain readily with phloxine and methylene blue and are easily observed by light microscopy (Fig. 5) (Bos, 1972; Verhoyen & Horvat, 1973; Bos et al., 1978a). In ultrathin sections, tubular and lamellar structures and pinwheels were observed (Horvat & Verhoyen, 1975a, 1975b; Paludan, 1980), characteristic of the potyvirus group.
Notes
Leek yellow stripe virus is distantly related to onion yellow dwarf virus (Bos, 1976) but, unlike it, does not infect Allium fistulosum, readily induces lesions in Chenopodium amaranticolor and C. quinoa, and is poorly infectious to onion and shallot; onion yellow dwarf virus infects leek only with difficulty. Yellow dwarf-affected crops of onion and shallot do not act as sources of virus for leek (Bos et al., 1978a) and these species seem to play little role in the ecology of leek yellow stripe virus: leek yellow stripe disease is prevalent only in areas with year-round cultivation of leek. Several publications that report the occurrence of onion yellow dwarf virus in leek, including those of Bremer (1937) and Kupke (1957), must have been dealing with leek yellow stripe virus.
Garlic yellow streak virus, reported from New Zealand (Mohamed & Young, 1981), is distantly related to leek yellow stripe and onion yellow dwarf viruses but does not infect leek or onion; it further differs from leek yellow stripe virus in failing to induce local lesions in C. amaranticolor and C. quinoa. The affinities of other potyviruses reported from garlic in France (Cadilhac et al., 1976), Venezuela (Lastra, Ladera & Debrot, 1979) and Japan (Lee et al., 1979) are not clear.
Other viruses reported naturally to infect leek are tomato black ring virus, a nepovirus (Calvert & Harrison, 1963; Graichen, 1975), and shallot latent virus, a carlavirus (Bos et al., 1978a; Bos, Huttinga & Maat, 1978b).
Figures

Leek (Allium porrum) with yellow stripe symptoms after natural infection.

Leek (Allium porrum) with yellow stripe symptoms after natural infection.

Leaf of Chenopodium amaranticolor with local lesions 39 days after inoculation.

Leaf of Chenopodium quinoa with local lesions 22 days after inoculation.

Inclusion bodies in epidermal cells of leek after staining with phloxine and methylene blue in Christie’s solution. i, inclusion bodies; n, nucleus. Bar represents 20 µm.
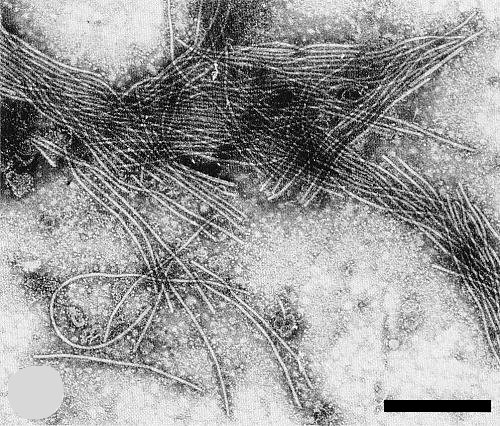
Electron micrograph of cluster of virus particles in crude sap from naturally infected leek plant. Bar represents 500 nm.
References list for DPV: Leek yellow stripe virus (240)
- Bos, Gewasbescherming 3: 81, 1972.
- Bos, CMI/AAB Descr. Pl. Viruses 158, 4 pp., 1976.
- Bos, Huijberts, Huttinga & Maat, Neth. J. Pl. Path. 84: 185, 1978a.
- Bos, Huttinga & Maat, Neth. J. Pl. Path. 84: 227, 1978b.
- Bremer, Phytopath Z. 10: 79, 1937.
- Cadilhac, Quiot, Marrou & Leroux, Annls Phytopath. 8: 65, 1976.
- Calvert & Harrison, Hort. Res. 2: 115, 1963.
- Graichen, Arch. Phytopath. PflSch. 11: 399, 1975.
- Graichen, Arch. Phytopath. PflSch. 14: 1, 1978.
- Horvat & Verhoyen, Parasitica 31: 55, 1975a.
- Horvat & Verhoyen, Phytopath. Z. 83: 328, 1975b.
- Huttinga, Neth. J. Pl. Path. 81: 81, 1975.
- Kupke, Rhein. Monatschr. Gem.- Obst.- u. Gartenb. 45: 173, 1957.
- Lastra, Ladera & Debrot, Phytopathology 69: 1036, 1979.
- Lee, Yamazaki, Osaki & Inouye, Ann. Phytopath. Soc. Japan 45: 727, 1979.
- Mohamed & Young, Ann. appl. Biol. 97: 65, 1981.
- Paludan, Tiddskr. PlAvl 84: 371, 1980.
- Verhoyen & Horvat, Parasitica 29: 16, 1973.