Details of DPV and References
DPV NO: 243 July 1981
Family: Caulimoviridae
Genus: Caulimovirus
Species: Cauliflower mosaic virus | Acronym: CaMV
This is a revised version of DPV 24
Cauliflower mosaic virus
R. J. Shepherd Dept. of Plant Pathology, University of California, Davis, CA 95616, USA
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
Described by Tompkins (1937).
- Selected synonyms
- Brassica virus 3 (Rev. appl. Mycol. 36: 303)
- Broccoli mosaic virus (Rev. appl. Mycol. 22: 122)
- Cabbage mosaic virus (Rev. appl. Mycol. 19: 65)
- Cabbage virus B (Rev. appl. Mycol. 24: 438)
- Broccoli mosaic virus (Rev. appl. Mycol. 22: 122)
A virus with small spherical particles about 50 nm in diameter containing a single molecule of circular double-stranded DNA. The DNA appears to replicate in the nuclei of infected cells as a plasmid. The particles are assembled in electron-dense inclusion bodies in the cytoplasm. The virus has a restricted host range and a world-wide distribution in temperate regions. It is transmitted by many aphid species in a non-persistent or semi-persistent manner.
Main Diseases
Induces mosaic and mottle diseases of many cruciferous crop plants and ornamental species, particularly the various cultivars of Brassica campestris and B. oleracea (Broadbent & Tinsley, 1953; Tompkins, 1937). Often found in mixed infections with turnip mosaic virus.
Geographical Distribution
World-wide in temperate regions.
Host Range and Symptomatology
With the exception of Nicotiana clevelandii (Hills & Campbell, 1968) and Datura stramonium (Lung & Pirone, 1972), only members of the Cruciferae have been reported as hosts. The virus is readily transmissible mechanically using an abrasive. Symptoms of the various strains differ considerably.
-
Diagnostic species
- Brassica oleracea
var. botrytis (cauliflower): Inoculated leaves usually symptomless; systemically infected leaves show initial vein-clearing, gradually replaced by green vein-banding (Fig. 1). Faint vein-clearing may persist in some varieties as the only chronic symptom (Broadbent & Tinsley, 1953). Even this symptom may disappear at high temperatures. - B. campestris (turnip or tendergreen mustard): Some inoculated leaves
develop chlorotic local lesions under cool conditions (Fig. 3); systemic symptoms
are vein-clearing, followed by chlorotic mottle (Fig. 2).
- Matthiola incana var. annua (annual stock): Systemic chlorotic mottle.
-
Propagation and assay species
- B. campestris cv. Just Right (a hybrid variety that gives uniform reactions). Some isolates give well-defined local lesions on this host that are useful for bioassay (Fig. 3) (Tomlinson & Shepherd, 1978).
Strains
Minor variants can be distinguished by their virulence in cauliflower or turnip (Broadbent & Tinsley, 1953). Severe strains cause stunting and death; mild ones induce almost no symptoms in chronic infections. Some atypical strains cause necrosis of leaf veinlets. In nature the virus usually occurs as a mixture of strains, some giving chlorotic lesions on Just Right turnip and others giving no local symptoms. Some strains are not transmissible by aphids (Lung & Pirone, 1973; 1974). A serological variant is known from Armoracia rusticana (horseradish).
Transmission by Vectors
At least 27 aphid spp. have been reported to transmit the virus (Kennedy, Day & Eastop, 1962) in a non-persistent or semi-persistent manner. All instars transmit and there is no latent period. Aphids can acquire the virus in 1-2 min and immediately thereafter can inoculate plants in less than 1 min. The virus is retained by aphids for periods of a few hours (van Hoof, 1954) or up to 3 days depending on the aphid species (Chalfant & Chapman, 1962). Transmission is relatively unaffected by post-acquisition feeding activity (Hamlyn, 1955). Isolates that are not transmissible by aphids become so if the aphids are first allowed to feed on plants infected with a transmissible isolate, or if the source plants are also infected with an aphid-transmissible isolate (Lung & Pirone, 1973; 1974). The transmission factor is probably a virus-induced, non-structural protein that functions in acquisition or inoculation of virus by the vector.
Transmission through Seed
None reported.
Serology
The virus is moderately immunogenic in rabbits. Virus particles diffuse slowly into 1% agar gels in double-diffusion tests and should be added 24 h before antiserum. The particles occur in plants in insufficient amounts or are released from inclusion bodies too slowly to give positive reactions in gel-diffusion or tube precipitin tests with plant extracts (Pirone, Pound & Shepherd, 1961). In immunosorbent tests, as little as 5 ng/ml can be detected with enzyme-linked antibody, or as little as 1 ng/ml with radioisotope-labelled antibody (Ghabrial & Shepherd, 1980). For immunosorbent tests, infected plant tissue should be homogenized in at least 50 vol. buffer or the extracts sonicated briefly to ensure the release of virus particles from the inclusion bodies.
Relationships
The virus is serologically related to dahlia mosaic virus (Brunt, 1966; 1971), carnation etched ring virus (Hollings & Stone, 1969), strawberry vein-banding virus (Morris et al., 1980), and perhaps to other caulimoviruses.
Stability in Sap
In cauliflower sap the thermal inactivation point is 75-80° C (10 min), dilution end-point c. 10-3 and longevity (at 20°C) 5-7 days.
Purification
Most investigators have used one of the following methods:
Pirone, Pound & Shepherd (1960). Homogenize tissue in 0.5 M phosphate (pH 7.5), add n-butanol to 8.5% (v/v) and centrifuge at 8000 g. Retain supernatant fluid. Concentrate the virus by 2 to 3 cycles of differential centrifugation, using water as the solvent; for large volumes of extract, the virus can first be precipitated by adding sodium chloride (to 0.05 M) and polyethylene glycol, M. Wt 6000 (100 g/l of extract), and then resuspending it in water (Shepherd, Bruening & Wakeman, 1970). Finally, purify the virus by density gradient centrifugation or by centrifugation to equilibrium in CsCl gradients (Shepherd, Wakeman & Romanko, 1968).
Hull, Shepherd & Harvey (1976), similar to the method of Gomec (1973) for dahlia mosaic virus. Grind chilled leaves (1 g/ml of buffer) in 0.5 M phosphate (pH 7) containing 0.75% sodium sulphite. Filter and add Triton X-100 to 2.5% (w/v) and urea to 1 M. Stir the homogenate overnight at 4°C and concentrate the virus by one cycle of differential centrifugation with resuspension in water. Purification is completed by rate zonal centrifugation in sucrose density gradients. The overnight incubation in Triton + urea is necessary with many strains to release the virus particles from the inclusion bodies. With some strains, such as CM4-184, the inclusion bodies break down rapidly after the addition of Triton + urea so that the homogenates can be fractionated immediately.
Gardner & Shepherd (1980) describe a procedure for rapid isolation of small quantities of virus DNA free of cellular DNA. Homogenize chilled tissue in 0.02 M Tris (pH 7.0), 0.02 M EDTA and 1.5 M urea using a Brinkman Polytron homogenizer. Add Triton X-100 to 2% (w/v) and subject the preparation to 1 cycle of differential centrifugation, resuspending the high speed pellets in 0.1 M Tris (pH 7.4) plus 2.5 mM MgCl2. Cellular DNA is then digested with DNase (10 µg/ml) at 37°C for 10 min. The virus is then treated with proteinase K in 1% sodium dodecyl sulphate at 65°C. After phenol extraction and precipitation with ethanol, the virus DNA (dissolved in 10 mM Tris + 0.1 mM EDTA, pH 8.0) is suitable for digestion with restriction endonucleases, or for molecular cloning. An amount of DNA equivalent to 20-40 mg of virus per kg of tissue can be obtained consistently by this procedure.
Properties of Particles
Not readily disrupted in any of the common denaturing protein solvents or chaotropic reagents unless the solutions are heated. Proteolysis with either Pronase or fungal proteinase K in the presence of dodecyl sulphate is effective in releasing the DNA (Shepherd et al., 1970; Shepherd, 1976). The particles sediment as a single component. Interparticle interactions significantly affect both sedimentation and diffusion coefficients in low ionic strength media down to about 0.1 mg/ml.
Sedimentation coefficient (s°20,w):208.2 (± 1.1) S at infinite dilution. The s°20,w increases by 8 S for each 1 mg/ml increase in virus concentration up to about 3 mg/ml (Hull et al., 1976).
Diffusion coefficient (D20,w):0.75 (± 0.04) x 10-7 cm2/sec with a marked concentration dependence in low ionic strength solvents (Hull et al., 1976).
Partial specific volume (v): 0.704 ± 0.007 g/ml over the measured concentration range of 1.2 to 0.12 mg/ml (Hull et al., 1976).
Molecular weight: 22.8 ± 1.4 x 106 calculated from the Svedberg equation.
Phosphorus content: 1.63%.
Particle Structure
The particles are approximately spherical but flatten considerably when preparations are air-dried on electron microscope grids (Pirone et al., 1961). In potassium phosphotungstate the particles have a diameter of c. 50 nm and an electron-dense centre of 20 nm diameter (Fig. 8). In ammonium molybdate or methylamine tungstate the diameter is 50.3 + 1.4 nm (Hull et al., 1976). In uranyl acetate the particles appear to be slightly smaller (c. 45 nm) with no electron-dense centre (Hills & Campbell, 1968). The hydrated diameter, calculated from the diffusion coefficient, is 57 nm (Hull et al., 1976).
Particle Composition
Nucleic acid: Double-stranded DNA, M. Wt about 5 x 106, about 17% of particle weight calculated from the phosphorus content (Hull et al., 1976) and nucleotide base ratios (Shepherd et al., 1970). G + C = 43%; Tm = 87.2°C in 0.15 M NaCl, 0.015 M citrate (pH 7.0) with hyperchromicity of 33-36%; buoyant density = 1.702 g/cm3 in CsCl; non-reactive to formaldehyde (Shepherd et al., 1970), contour length of 2.31 µm (Shepherd & Wakeman, 1971) or 2.47µm (Russell et al., 1971). Most preparations contain linear and circular forms as revealed by electron microscopy, as well as two sedimenting forms (17.1 and 19.0 S), and two components in gel electrophoresis. Only the circular form is infective (Hull & Shepherd, 1977); the linear molecule is probably a breakage product of the circular form (Hull & Howell, 1978; Volovitch, Drugeon & Yot, 1978). Viral DNA that has been cloned in E. coli is infective for plants (Howell, Walker & Dudley, 1980). The complete sequence of about 8000 nucleotide pairs has been determined for virus DNA taken directly from virus particles (Franck et al., 1980) or after molecular cloning in Escherichia coli (Gardner et al., 1981).
Protein: Coat protein can be isolated by degradation of virus in hot 6 M guanidine HCl and passage through a column of Biogel P-300. The DNA emerges in the void volume. Gel electrophoresis of preparations made by boiling virus in 3.5% sodium dodecyl sulphate reveals several proteins; the major components have M. Wt (x 10-3) of 32-34, 37-39, 40-44 and 64-70, and may be accompanied by a variety of minor components (Tezuka & Taniguchi, 1972; Kelly, Cooper & Walkey, 1974; Brunt et al., 1975; Hull & Shepherd, 1975). There is evidence that the major components are either degradation or aggregation products of a single protein (Al Ani, Pfeiffer & Lebeurier, 1979), which has a M. Wt of either 44,000 or 58,000 and is phosphorylated (Hahn & Shepherd, 1980). The coat protein is basic with a high content of lysine and arginine (Brunt et al., 1975). The nucleotide sequence of the DNA suggests that some of the other gene products are also very basic in nature (Gardner et al., 1981).
Other constituents: None reported. The particles contain less than 0.1% fatty acids in gas chromatographic analyses for triglycerides and phospholipids (Hull et al., 1976).
Genome Properties
The viral genome has been mapped physically with the aid of restriction endonucleases (Meagher, Shepherd & Boyer, 1977; Volovitch et al., 1978; Howarth et al., 1981; Hull & Howell, 1978; Lebeurier et al., 1978; Volovitch et al., 1979; Hull, 1980; Gardner et al., 1980). The dsDNA is a relaxed molecule (i.e. not super-coiled) and has interruptions at specific sites, two in one strand and one in the other (Volovitch et al., 1978) (Fig. 9). Interruptions consist of a break in one strand of the polynucleotide chain with a short extension of redundant sequences to produce an overlap of the double helix. This protrudes as a short single-stranded extension probably of the 5'-end (Franck et al., 1980). The strand with a single interruption, termed the a-strand, contains numerous nonsense codons, whereas the complementary strand has six long open-reading regions (Fig. 9); this is consistent with the finding that only the a-strand is transcribed to RNA (Howell & Hull, 1978; Hull et al., 1979; Volovitch et al., 1980; Guilfoyle, 1980). Plant RNA polymerase II appears to be active in this transcription (Guilfoyle, 1980). The open-reading regions probably correspond directly to amino acid sequences comprising virus proteins in a co-linear fashion (Franck et al., 1980). Open-reading region II (500 base pairs) can be deleted without much loss of infectivity. Comparison of the DNA sequences of strain CM-1841 and a closely related deletion mutant, strain CM4-184, both defective in aphid transmission, shows that 421 base pairs have been deleted in the latter (Howarth et al., 1981).
Complete transcriptional maps of the virus DNA are not yet available but a major (19 S) RNA transcript, termed P66, has been isolated and shown to be translated in vitro to yield a non-structural protein of 66,000 M. Wt (Odell & Howell, 1980; Al Ani et al., 1980) which may be a component of the cytoplasmic inclusion bodies. The mapping coordinates for this RNA largely correspond to open-reading region 6, ending near the single-stranded break (overlap) in the a-strand of the DNA; its 3'-end is polyadenylated (Odell & Howell, 1980). The major protein constituent of the inclusion body, the 55,000 M. Wt ‘matrix protein’, is host-specified (R. J. Shepherd, R. Richins & S. D. Daubert, unpublished data). Highly radioactive, unit-length viral DNA can be prepared from isolated plant nuclei to which radioactive deoxyribonucleoside triphosphates have been administered (O. Ansa, V. W. Bowyer & R. J. Shepherd, unpublished data). No virus can be detected in such nuclei, and hence it seems probable that viral DNA replicates in cell nuclei as a plasmid.
Relations with Cells and Tissues
A unique type of inclusion body occurs in the cytoplasm of cells infected with caulimoviruses (Fig. 4, Fig. 5, Fig. 7) (Fujisawa et al., 1967). These are a conspicuous feature of stripped epidermis stained with 1% phloxine and viewed with the light microscope (Fig. 5). These inclusion bodies consist of an electron-dense matrix in which virus particles are embedded (Fig. 4). Most of the virus particles that appear in cells seem to be associated with the inclusion bodies. Though the inclusion bodies differ in size with virus strain (compare Fig. 4 and Fig. 7) (Shalla, Shepherd & Peterson, 1980), they may exceed 20 µm in diameter (Mamula & Milicic, 1968). They start as granular electron-dense patches in the cytoplasm and appear to grow by accretion throughout the course of infection. The ‘matrix protein’ makes up the bulk of these inclusions and accounts for the intensity of their staining with osmium (Shepherd, Richins & Shalla, 1980). Virus particles appear in the interior of the bodies, either embedded in the matrix or filling the vacuoles as if they are assembled within the inclusions. A modified form of the inclusion can be isolated as an uniformly electron-dense granule (Fig. 6) (Shepherd et al., 1980). Although considerable amounts of transcribed viral RNA can occur in the nucleus (Guilfoyle, 1980), no virus particles are found there. Free virus particles occur in small amounts in the cytoplasm near the inclusions and infrequently in plasmodesmata. Cell wall protrusions associated with masses of vesicles and convoluted tubules are common in infected tissues (Conti et al., 1972).
Notes
The virus differs from other non-persistent, aphid-borne viruses found in crucifers in its relatively high thermal inactivation point (75-80°C), its greater longevity in vitro (5-7 days), its particle size and its restricted host range (Walker, LeBeau & Pound, 1945). Unlike turnip mosaic virus, with which it is commonly found in mixed infections, cauliflower mosaic virus does not infect species of Solanaceae (with the exception of Nicotiana clevelandii and Datura stramonium) or Chenopodiaceae. Its longer retention by feeding aphids provides a useful way of separating it from turnip mosaic virus. The characteristic inclusion bodies it induces in infected plants provide an additional means of diagnosis. Even more convenient and specific for identification, however, are the sensitive enzyme-linked or radioisotope-immunosorbent assays (Ghabrial & Shepherd, 1980).
Figures

A leaf of cauliflower (Brassica oleracea) infected with cauliflower mosaic virus showing chronic green vein-banding symptoms (photo courtesy of J A. Tomlinson).

Mottle and generalized chlorosis in a leaf of turnip (B. campestris cv. Just Right) infected with strain CM4-184, a relatively mild strain that produces good virus yields and large inclusion bodies but causes little inhibition of growth of most varieties of B. campestris.
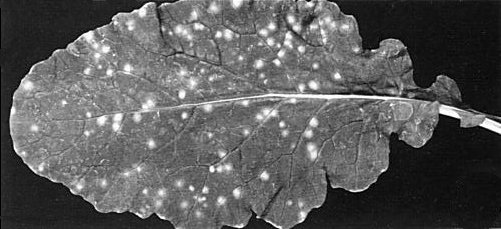
Chlorotic local lesions on Just Right turnip inoculated with a selected variant of the cabbage B strain. The plant was kept at 16°C with a 16 h daylength for 20 days after inoculation.

Section of an inclusion body in the cytoplasm of a cell of B. campestris infected with the Campbell strain. Note the location in the cytoplasm, the electron-dense matrix with embedded virus particles and the lack of an external membrane.

Inclusion bodies (arrows) in the epidermis stripped from infected B. campestris. The tissue was stained with 1% phloxine for a few minutes and then repeatedly rinsed in saline.

Isolated inclusion bodies showing (left) the native, vacuolated type with occluded virus particles and (right) the granular type, which consists solely of the 55,000 dalton matrix protein (Shepherd et al., 1980).

Section of an inclusion body in B. campestris infected with the Bari strain. Note the less well developed inclusion body with only a small amount of matrix material (arrow).
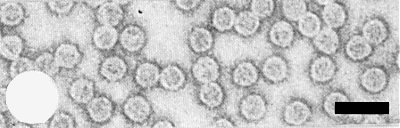
Virus particles of strain CM4-184. Bar represents 100 nm.
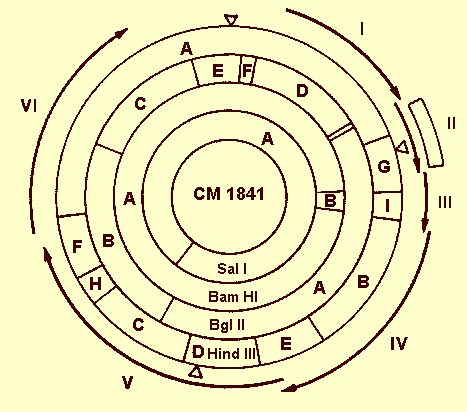
A physical map of the circular genome of strain CM1841 showing the sites of cleavage by Sal I, Bam HI, Bgl II, and Hin dill restriction endonucleases. The open-reading regions from the DNA sequence are indicated by peripheral arrowed lines I to VI. The deletion of most of region II of homologous strain CM4-184 is indicated by the box. The three single-stranded interruptions are indicated by open triangles. (Diagram taken from Howarth et al., 1981).
References list for DPV: Cauliflower mosaic virus (243)
- Al Ani, Pfeiffer & Lebeurier, Virology 93: 188, 1979.
- Al Ani, Pfeiffer, Whitechurch, Lesot, Lebeurier & Hirth, Annls Virol. (Inst. Pasteur) 131E: 33, 1980.
- Broadbent & Tinsley, Pl. Path. 2: 88, 1953.
- Brunt, Virology 28: 778, 1966.
- Brunt Ann. appl. Biol. 67: 357, 1971.
- Brunt, Barton, Tremaine & Stace-Smith, J. gen. Virol. 27: 101, 1975.
- Chalfant & Chapman, J. econ. Entomol. 55: 584, 1962.
- Conti, Vegetti, Bassi & Favali, Virology 47: 644, 1972.
- Franck, Guilley, Jonard, Richards & Hirth, Cell 21: 285, 1980.
- Fujisawa, Rubio-Huertos, Matsui & Yamaguchi, Phytopathology 57: 1130, 1967.
- Gardner & Shepherd, Virology 106: 159, 1980.
- Gardner, Melcher, Shockey & Essenberg, Virology 103: 250, 1980.
- Gardner, Howarth, Hahn, Brown-Luedi, Shepherd & Messing, Nucl. Acids Res. 9: 2871, 1981.
- Ghabrial & Shepherd, J. gen. Virol. 48: 311, 1980.
- Gomec, Ph.D Thesis, Univ. of California, Davis, 1973.
- Guilfoyle,Virology 107: 71, 1980.
- Hahn & Shepherd, Virology 107: 295, 1980.
- Hamlyn, Pl. Path. 4: 13, 1955.
- Hills & Campbell, J. Ultrastruct. Res. 24: 134, 1968.
- Hollings & Stone, Rep. Glasshouse Crops Res. Inst., 1968: 102, 1969.
- Howarth, Gardner, Messing & Shepherd, Virology 112: 678, 1981.
- Howell & Hull, Virology 86: 468, 1978.
- Howell, Walker & Dudley, Science N. Y. 208: 1265, 1980.
- Hull, Virology 100: 76, 1980.
- Hull & Howell, Virology 86: 482, 1978.
- Hull & Shepherd, Virology 70: 217, 1975.
- Hull & Shepherd, Virology 79: 216, 1977.
- Hull, Shepherd & Harvey, J. gen. Virol. 31: 93, 1976.
- Hull, Covey, Stanley & Davies, Nucl. Acids Res. 7: 669, 1979.
- Kelly, Cooper & Walkey, Microbios 10: 239, 1974.
- Kennedy, Day & Eastop, A conspectus of aphids as vectors of plant viruses, London, Commonwealth Institute of Entomology, 114 pp., 1962.
- Lebeurier, Whitechurch, Lesot & Hirth, Gene 4: 213, 1978.
- Lung & Pirone, Phytopathology 62: 1473, 1972.
- Lung & Pirone, Phytopathology 63: 910, 1973.
- Lung & Pirone, Virology 60: 260, 1974.
- Mamula & Milicic, Phytopath. Z. 61: 232, 1968.
- Morris, Mullin, Schlegel, Cole & Alosi, Phytopathology 70: 156, 1980.
- Meagher, Shepherd & Boyer, Virology 80: 362, 1977.
- Odell & Howell, Virology 102: 349, 1980.
- Pirone, Pound & Shepherd, Nature, Lond., 186: 656, 1960.
- Pirone, Pound & Shepherd, Phytopathology 51: 541, 1961.
- Russell, Follett, Subak-Sharpe & Harrison, J. gen. Virol. 11: 129, 1971.
- Shalla, Shepherd & Peterson, Virology 102: 381, 1980.
- Shepherd, Adv. Virus Res. 20: 305, 1976.
- Shepherd & Wakeman, Phytopathology 61: 188, 1971.
- Shepherd, Wakeman & Romanko, Virology 36: 150, 1968.
- Shepherd, Bruening & Wakeman, Virology 41: 339, 1970.
- Shepherd, Richins & Shalla, Virology 102: 389, 1980.
- Tezuka & Taniguchi, Virology 47: 142, 1972.
- Tompkins, J. agric. Res. 55: 33, 1937.
- Tomlinson & Shepherd, Ann. appl. Biol. 90: 223, 1978.
- van Hoof, Tijdschr. PlZiekt. 60: 267, 1954.
- Volovitch, Drugeon & Yot, Nucl. Acids Res. 5: 2913, 1978.
- Volovitch, Drugeon, Dumas, Haenni & Yot, Eur. J. Biochem. 100: 245, 1979.
- Volovitch, Chouikh, Kondo & Yot, FEBS Lett. 116: 257, 1980.
- Walker, LeBeau & Pound, J. agric. Res. 70: 379, 1945.