Details of DPV and References
DPV NO: 251 July 1982
Family: Rhabdoviridae
Genus: Nucleorhabdovirus
Species: Cereal chlorotic mottle virus | Acronym: CCMoV
Cereal chlorotic mottle virus
R. S. Greber Department of Primary Industries, Indooroopilly, 4068, Australia
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
Described by Greber (1977; 1979) and Greber & Gowanlock (1979).
A virus with bacilliform particles c. 65 x 240 nm containing RNA, protein and, probably, lipid. Transmitted in a persistent manner by cicadellid leafhoppers but not by inoculation of sap. Infects plants only in the family Gramineae; natural infection occurs in maize and weed grasses and less frequently in temperate cereals. Reported only from eastern Australia.
Main Diseases
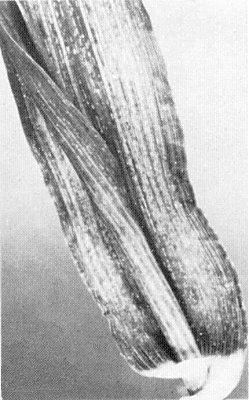
In maize, the virus causes fine chlorotic striations in the leaves (Fig. 1), with variable stunting and tassel sterility (Greber, 1979; 1981); epiphytotics occur sporadically, occasionally reaching 60% incidence but usually 0-10%. Naturally infected temperate cereals, e.g. barley (Hordeum vulgare; Fig. 2) and wheat (Triticum aestivum; Fig. 3) also show striations and stunting but epiphytotics do not occur in winter-grown crops. Species in several grass genera provide a natural reservoir of virus (Greber, 1981).
Geographical Distribution
Reported only from Eastern Australia (latitude 19°-29°S) but range may be wider.
Host Range and Symptomatology
Known plant hosts are all in the family Gramineae, scattered through the sub-families, but with fewer hosts in the Andropogonoidae. The following species are hosts: Avena sativa, Bromus unioloides, Digitaria ciliaris, Dinebra retroflexa, Echinochloa colona, Eleusine indica, Eragrostis cilianensis, Hordeum vulgare, Leptochloa filiformis, Secale cereale, Setaria italica, Setaria verticillata, x Triticosecale (Triticum aestivum x Secale cereale), T. aestivum, T. monococcum, T. tauschii, Urochloa panicoides, Zea mays. Some of these species (e.g. Dinebra retroflexa and Triticum spp.) are more susceptible than others. No marked genotype differences have been noted within species except in maize (Greber, 1981).
This virus is not transmitted by inoculation of sap, only by the leafhopper vectors; young seedlings are the most readily infected. Symptoms in most hosts consist of various types of chlorotic striation with or without stunting. Striations tend to decrease in intensity in chronic infections. Some degree of sterility is usually caused, particularly in the male inflorescence in maize.
-
Diagnostic species
- Zea mays
(maize). Some lines and hybrids are highly resistant (Greber, 1981), others are highly susceptible and these can be used as differential hosts. Susceptible lines (e.g. B37, H84) produce fine short chlorotic striations (Fig. 1) along the veins over the leaf lamina, leaf sheath and ear husks. Symptoms appear c. 14 days after leafhopper inoculation at 25°C. - Triticum spp. (wheat) and x Triticosecale. First symptoms are
prominent chlorotic striations (Fig. 3) which take up much of the area of the
first few leaves emerging after inoculation. Striations in later leaves are
usually finer and less prominent.
- Dinebra retroflexa. Readily infected and shows persisting chlorotic striations.
- Leptochloa filiformis. Severe chlorotic striations which turn to rusty brown necrosis. The severe leaf symptoms are accompanied by deformation of the inflorescence.
- Sorghum bicolor (sorghum) and Oryza sativa (rice) are not infected.
- Dinebra retroflexa. Readily infected and shows persisting chlorotic striations.
-
Propagation species.
- Maize is the best source of virus for purification. Cultures can be maintained in Dinebra retroflexa. Vectors can be reared free of the virus on Chioris gayana (for Nesoclutha pallida) and Sorghum bicolor (for Cicadulina bimaculata).
-
Assay species
- Dinebra retroflexa
is a good test plant in leafhopper transmission experiments.
Strains
No evidence available of strain differentiation.
Transmission by Vectors
Efficiently transmitted by at least two species of cicadellid leafhopper, Nesoclutha pallida (Greber, 1977; 1979) and Cicadulina bimaculata (Greber, 1981). The virus was transmitted efficiently by leafhoppers that were injected with a preparation of purified virus but not by insects that fed on the same preparation through a membrane (R. Greber, unpublished data). For N. pallida, the minimum acquisition access period was 4 h (R. Greber, unpublished data) and the latent period was 12-27 days for insects that acquired the virus from plants (Greber, 1979) or 8-10 days for insects injected with purified virus preparations (R. Greber, unpublished data). The virus usually persists in the vector for life but some insects cease to transmit up to 4 weeks before death (Greber, 1979). The virus is acquired both by adults and by nymphs and is transmitted through the moult (Greber, 1979). The virus infects cells of brain and salivary glands of both vector species (Fig. 8, Fig. 9) and apparently multiplies in the vector (Greber & Gowanlock, 1979 and unpublished data). The vectors have a wide feeding range among species of Gramineae but acquisition from and transmission to some host species is effected only by C. bimaculata, though N. pallida injected with infective sap from these species will transmit to other species (Greber, 1981).
Transmission through Seed
Not transmitted through seed of maize or D. retroflexa.
Serology
Weakly to moderately immunogenic. Antisera prepared by intramuscular injection of purified virus had titres of 1/32 to 1/128 in gel-diffusion tests. Two, sometimes three, precipitin lines are formed (Fig. 10) and the relative position and intensity of these lines varies in different tests. Addition of 1% Triton X-100 to the antigen well usually enhances the reaction. Virus also reacts in ring-precipitin tests, and the gel-diffusion reaction with infective sap is useful in diagnosis. All major antigens are sedimented from fresh infective sap (100,000 g, 110 min) but one, presumably the G-protein, is released as a soluble antigen after treatment with 1% Triton X-100 (Fig. 10) or by degradation of stored preparations.
Relationships
Particle morphology and virus/vector relations place the virus in the plant rhabdovirus group (Matthews, 1979). It is classified within Subgroup II (Peters, 1981) because of the substantial association of the particles with the nuclei of infected plant and insect cells (Greber & Gowanlock, 1979). The rhabdoviruses infecting Gramineae are transmitted either by delphacid or cicadellid leafhoppers but never by vectors of both types. This allows differentiation on both the type of vector and association of the particles with the nucleus. The virus resembles wheat striate mosaic virus in its cellular distribution and site of maturation and in having a cicadellid vector; however, no serological relationship has been demonstrated (Greber & Gowanlock, 1979; R. G. Timian, personal communication). No serological reaction was obtained with antiserum to maize mosaic virus (Greber & Gowanlock, 1979), a virus which also differs in having a delphacid vector and in not being reported to infect festucoid grasses. Barley yellow striate mosaic virus and northern cereal mosaic virus are delphacid-transmitted and have characteristics of Subgroup I (Peters, 1981).
Stability in Sap
Virus in Dinebra retroflexa sap, assayed by injection into Nesoclutha pallida, had a thermal inactivation point (10 min) between 45 and 50°C. Purified virus, assayed in the same way was infective after 7 days at 4°C (R. Greber, unpublished data).
Purification
A modification of the method of Jackson & Christie (1977) is satisfactory. Tissue (40-80 g) with good symptoms is checked for high particle concentration by negative stain electron microscopy and then ground in a cold mortar, squeezed through cloth and re-extracted using 0.1 M Tris-HCl buffer, pH 8.4, containing 0.04 M sodium sulphite. The metal salts used by Jackson & Christie (1977) are omitted. After a low speed centrifugation and adjusting to pH 7.4 the extract is centrifuged in tubes containing solutions of 300 mg/ml sucrose layered over 600 mg/ml sucrose; the virus is collected from the interface and filtered through Celite (10 g in a 9 cm funnel). The filtrate and washings are then passed through two successive gradients of 100-400 mg/ml sucrose and 300-600 mg/ml sucrose and pelleted in the manner described by Jackson & Christie (1977). Concentrated preparations have a milky appearance and gradient bands are very easily located. Most particles from purified preparations appear relatively intact when fixed with glutaraldehyde before electron microscope examination.
Properties of Particles
The particles sediment as a single band with a buoyant density in sucrose solution of 1.165 g/cm3. The particle mass (from density and dimensions) is about 5.7 x 108 daltons (R. Greber, unpublished data). Light-scattering is substantial, but the uncorrected A260/A280 ratio is 1.17. There is a gradual increase of uncorrected absorbance from A280 through to A240.
Particle Structure
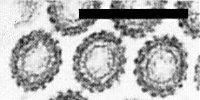
Normally bacilliform, 65 nm x 240 nm in thin section and 75 nm x 250 nm when fixed in glutaraldehyde before negative staining (Fig. 5, top particle). Unfixed particles disrupt in phosphotungstate negative stain to yield bullet-shaped structures, the larger particles measuring 75 nm x 215 nm (Fig. 5, second particle). A prominent net-like surface pattern (Fig. 6) is evident both in negative stain and in tangential thin sections (Greber, 1979; Greber & Gowanlock, 1979). This pattern correlates with the number of projections seen around the perimeter (52 in longitudinal sections and 17 in cross sections; Fig. 7). The helix, which is infrequently penetrated by negative stain (Fig. 5, third particle), has c. 48 turns, approximately two turns for each row of projections. In cross section (Fig. 7) the densely stained, spiked outer zone is clearly separated from an inner stained circle; the central core (35 nm) is largely unstained, with no central dot. The envelope projections and net-pattern continue round the hemispherical ends, but at one end of each particle (at the point of detachment from the nuclear membrane) the inner structure is less densely stained within the hemisphere (Fig. 5, bottom particle).
Particle Composition
Nucleic acid: RNA (R. Greber, unpublished data), probably single-stranded negative sense, as in other rhabdoviruses (Peters, 1981), and arranged in a helix c. 5.5 µm in total length.
Protein: Five polypeptides are resolved by electrophoresis in polyacrylamide/SDS gels (J. Dale & R. Greber, unpublished data) with M. Wt. (x 10-3) of 77.5, 52.5, 46.5, 32 and 29; these probably correspond to proteins G, N, NS, M1 and M2 of other rhabdoviruses (Peters, 1981).
Relations with Cells and Tissues
Virus particles are present in most tissues of infected maize plants and in brain and salivary gland tissue of infective leafhoppers. The particles appear to receive their outer envelope at the inner nuclear membrane and are frequently arranged as a palisade of bullet-shaped structures projecting into the perinuclear space; they accumulate as bacilliform particles within the perinuclear space and in cytoplasmic vesicles (Fig. 4). In insect tissue, particles may also be embedded in granular viroplasm-like structures, within and around the nucleus. In some plant and vector cells, large accumulations of particles may form and may appear to be scattered throughout the nucleus. Unenveloped particles have not been observed, but isolated complete particles are often found surrounded by individual vesicle membranes.
Notes
The main economic host is maize. Cereal chlorotic mottle virus is cicadellid-transmitted, unlike maize mosaic virus (Herold, 1972), a member of plant rhabdovirus Sub-group II (Peters, 1981) and maize sterile stunt virus (Greber, 1982), a member of plant rhabdovirus Subgroup I, both of which have delphacid planthopper vectors. Particles of maize sterile stunt virus are readily distinguished from those of cereal chlorotic mottle virus in negatively stained leaf extracts (Greber, 1979), because they lack the prominent surface pattern and are longer and narrower (Fig. 6), and the stain penetration of the core is only half as wide.
Cereal chlorotic mottle virus resembles American wheat striate mosaic virus (Sinha & Behki, 1972; Jackson, Milbrath & Jedlinski, 1981) in several respects. Particles of both viruses accumulate mainly in the perinuclear space, both have cicadellid leafhopper vectors and the host range has a substantial coincidence. However, no serological relationship has been detected between the two viruses, and wheat striate mosaic virus differs from cereal chlorotic mottle virus in having a higher buoyant density in sucrose (1.22 g/cm3: Sinha, Harwalker & Behki, 1976), in having only one M-protein (Trefzer-Stevens & Lee, 1977), in not showing a prominent net-like surface pattern in negative stain, and in yielding a large amount of soluble antigen (Thottappily & Sinha, 1973).
Figures

Chlorotic striations caused by natural infection in maize.
Striation symptoms in naturally infected barley.
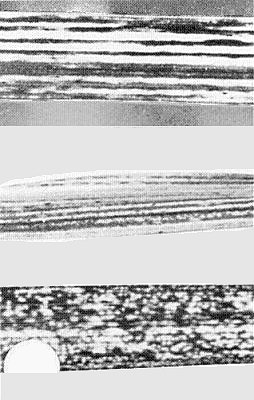
(From top): symptoms in wheat, Setaria italica and Eleusine coracana.

Thin section of infected maize cell showing perinuclear distribution of virus particles (V). Bar represents 1 µm.

Virus particles (from top): bacilliform, glutaraldehyde-fixed in negative stain; bullet-shaped, unfixed, in negative stain; helix penetration; longitudinal thin section showing projections and one end less densely stained (fixed with glutaraldehyde then osmium tetroxide, embedded in Spurr’s medium, stained with uranyl acetate and lead citrate). Bar represents 100 nm.

Prominent surface pattern and wider core penetration of cereal chlorotic mottle virus (three upper particles) compared in negative stain to maize sterile stunt virus (lower particle).
Transverse thin section of virus particles, showing protrusions. Bar represents 100 nm.
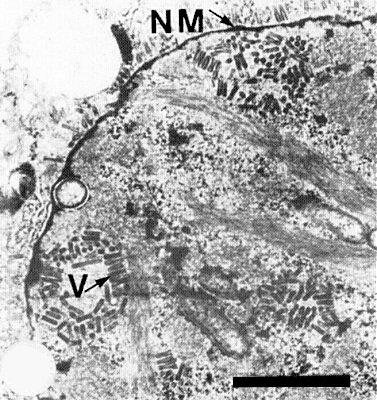
Thin section of infected salivary gland cell from Cicadulina bimaculata. Virus particles (V); nuclear membrane (NM). Bar represents 1 µm.

Thin section of infected brain cell from Nesoclutha pallida. Virus particles (V); nuclear membrane (NM). Bar represents 500 nm.

Gel-diffusion serological reactions with cereal chlorotic mottle virus antiserum (As). Top-purified virus (V), purified virus treated with Triton X-100 (VX), and infective sap (IX). Below-separation of antigens (p and s) by detergent treatment and differential centrifugation. Infective sap (IS). All other antigen wells filled with preparations treated with 1% Triton X-100: pellet 100,000 g, 110 min (P); supernatant fluid 100,000 g, 110 min (5); supernatant fluid 100,000 g, 20 min (S'); infective sap (IX).
References list for DPV: Cereal chlorotic mottle virus (251)
- Greber, Newsl. Aust. Pl. Path. Soc. 6: 17, 1977.
- Greber, Aust. J. agric. Res. 30: 433, 1979.
- Greber, Australas. Pl. Path. 10: 29, 1981.
- Greber, Aust. J. agric. Res. 33: 13, 1982.
- Greber & Gowanlock, Aust. J. biol. Sci. 32: 399, 1979.
- Herold, CMI/AAB Descr. Pl. Viruses 94, 4 pp., 1972.
- Jackson & Christie, Virology 77: 344, 1977.
- Jackson, Milbrath & Jedlinski, in Virus and Viruslike Diseases of Maize in the United States, pp. 51-76, ed. D. Gordon, J. Knoke & G. Scott, Ohio Agric. Res. Dev. Cent., Wooster, Ohio, 210 pp., 1981.
- Matthews, Intervirology 12: 132, 1979.
- Peters, CMI/AAB Descr. Pl. Viruses 244, 6 pp., 1981.
- Sinha & Behki, CMI/AAB Descr. Pl. Viruses 99, 4 pp., 1972.
- Sinha, Harwalkar & Behki, Phytopath. Z. 87: 314, 1976.
- Thottappilly & Sinha, Virology 53: 312, 1973.
- Trefzer-Stevens & Lee, Virology 78: 144, 1977.