Details of DPV and References
DPV NO: 252 July 1982
Family: Tombusviridae
Genus: Gallantivirus
Species: Galinsoga mosaic virus | Acronym: GaMV
Galinsoga mosaic virus
G. M. Behncken Department of Primary Industry, Meiers Road, Indooroopilly, Queensland, 4068, Australia
R. I. B. Francki Waite Agricultural Research Institute, University of Adelaide, Glen Osmond, South Australia, 5064, Australia
A. J. Gibbs Research School of Biological Sciences, Australian National University, Canberra, A.C.T., 2601, Australia
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
Described by
Behncken (1970).
A virus with isometric particles c. 34 nm in diameter which contain single-stranded RNA and sediment as a single component. Found only in Galinsoga parviflora (Compositae) in Queensland, Australia. It is readily transmitted by inoculation with sap. In naturally infected plants it is most concentrated in the roots and is transmitted through soil, apparently without the aid of a vector.
Main Diseases
The virus is found in nature only in potato weed, Galinsoga parviflora (Compositae), in which it is usually restricted to the roots. Of no known economic importance.
Geographical Distribution
Queensland, Australia.
Host Range and Symptomatology
The virus has a wide experimental host range, infecting species in seven dicotyledonous families, but in most is confined to the inoculated leaves. Readily transmitted by inoculation with sap.
-
Diagnostic species
- Chenopodium amaranticolor.
Small necrotic local lesions with chlorotic haloes (Fig. 4); no systemic infection. - C. quinoa. Necrotic local lesions; no systemic infection.
- Galinsoga parviflora. Chlorotic/necrotic local lesions; systemic vein clearing (Fig. 2) followed by a severe chlorotic mosaic (Fig. 3). Systemically infected leaves later develop areas of necrosis and become distorted, and the plant becomes stunted (Fig. 1).
- Phaseolus vulgaris cvs Bountiful, Pinto. Small reddish-brown necrotic local lesions; no systemic infection (Fig. 5).
- Spinacia oleracea. Systemic chlorotic spots and vein necrosis. Leaves produced later are symptomless.
- Galinsoga parviflora. Chlorotic/necrotic local lesions; systemic vein clearing (Fig. 2) followed by a severe chlorotic mosaic (Fig. 3). Systemically infected leaves later develop areas of necrosis and become distorted, and the plant becomes stunted (Fig. 1).
-
Propagation species
- Leaves of C. amaranticolor and P. vulgaris (cvs Hawkesbury Wonder, Canadian Wonder or Royal Windsor) are suitable sources for virus purification.
-
Assay species
- C. amaranticolor, C. quinoa
or P. vulgaris are satisfactory local lesion hosts.
Strains
None reported.
Transmission by Vectors
No vectors reported. The aphids Aphis gossypii and Myzus persicae failed to transmit the virus (Behncken, 1970). Transmission tests with the fungus Olpidium brassicae from roots of infected plants were also negative (Behncken, 1970; Shukla et al., 1979). The roots of G. parviflora plants became infected when grown (a) in naturally infective soil, (b) in autoclaved potting mixture to which infective leaf extract was added or (c) in the same pot of autoclaved soil as a leaf-inoculated plant, even when the tops of the two plants were separated by a glass partition (Shukla et al., 1979). These results suggest that the virus infects G. parviflora roots without the aid of a vector.
Transmission through Seed
The virus was not transmitted in seed of G. parviflora (Shukla et al., 1979).
Serology
An antiserum with a titre of 1/1024 in gel diffusion tests was readily obtained (Behncken, 1970).
Relationships
The virus shares properties with several viruses that have isometric particles sedimenting as one component, but its particles are serologically unrelated to those of any such virus with which it has been compared. In gel diffusion tests, galinsoga mosaic virus particles and/or antiserum failed to react with those of 19 viruses from nine taxonomic groups including red clover necrotic mottle (dianthovirus group), southern bean mosaic (sobemovirus group) and tomato bushy stunt and glycine mottle (tombusvirus group). Galinsoga mosaic virus particles also failed to react with antisera to the following ungrouped viruses: carnation mottle, hibiscus chlorotic ringspot, sowbane mosaic, tobacco necrosis, velvet tobacco mottle and uncharacterized viruses from Glycine max (Behncken, 1972) and Malvastrum coromandelianum (G. M. Behncken, unpublished data). Moreover its possession of a single genomic RNA species distinguishes galinsoga mosaic virus from red clover necrotic mottle and velvet tobacco mottle viruses, which have bipartite RNA genomes. The particles of galinsoga mosaic virus are larger than those of southern bean mosaic and velvet tobacco mottle viruses, are less angular in outline, and show some surface structure. Galinsoga mosaic virus shares most properties with tomato bushy stunt virus, which has particles of similar size, appearance and composition but with a larger sedimentation coefficient (T. Hatta, R. I. B. Francki & C. J. Grivell, unpublished data). Cells infected with either of these viruses contain multivesicular bodies which are of similar appearance but possibly develop from different organelles (Russo & Martelli, 1972; Appiano, Pennazio & Redolfi, 1978; T. Hatta, R. I. B. Francki & C. J. Grivell, unpublished data).
Stability in Sap
Sap from infected C. amaranticolor leaves lost infectivity after 10 min at 75-80°C, and when diluted to more than 10-6. At 25°C sap retained infectivity for 6 weeks.
Purification
Particles are readily purified from the inoculated leaves of either C. amaranticolor or bean. Leaves are ground in 0.1 M phosphate buffer (pH 7.0) containing 0.1% sodium sulphite, the extract clarified by homogenizing with chloroform-butanol, and the particles concentrated and purified by differential centrifugation.
Properties of Particles
In sucrose density gradients and during analytical centrifugation, the virus particles usually sediment as a single component.
Sedimentation coefficient (s20, w): 117.9±2.1 S; some preparations also contain a minor 48 S component.
A260/A280: 1.66. The 48 S component has a smaller absorbance ratio (Skotnicki & Gibbs, 1981).
The particles are stable in 1 M CaCl2 (Behncken, 1970).
Particle Structure
Particles are isometric, c. 34 nm in diameter with a rounded profile when negatively stained in uranyl acetate (Fig. 6). They have a verrucose surface but the arrangement of the subunits is not clear (T. Hatta, R. I. B. Francki & C. J. Grivell, unpublished data).
Particle Composition
Nucleic acid: RNA, probably single-stranded, infective when deproteinized. A single RNA species, of M. Wt c. 1.55 x 106, estimated by polyacrylamide gel electrophoresis under non-denaturing conditions. It constitutes about 22% of the particle, and its percentage base composition is G 24.8±0.3; A 28.1±0.6; C 21.8±1.0; U 25.3±0.7 (Skotnicki & Gibbs, 1981).
Protein: Coat protein is a single species of M. Wt c. 36,400, estimated by polyacrylamide gel electrophoresis. Percentage molar amino acid composition (and ‘best fitting’ integral number of amino acids); ala 7.07 (26), arg 4.44 (16), asx 9.30 (34), cys 1.61 (6), glx 8.08 (29), gly 9.97 (36), his 2.14 (8), ile 2.99 (11), leu 8.75 (32), lys 4.70 (17), met 0 (0), phe 9.30 (34), pro 4.47 (16), ser 8.89 (32), thr 9.09 (33), trp 0.81 (3), tyr 1.98 (7), val 6.42 (23); estimated total number of amino acids 363, M. Wt 39,505 (Skotnicki & Gibbs, 1981).
Relations with Cells and Tissues
Infected P. vulgaris or G. parviflora leaf cells have particles throughout their cytoplasm and in some vacuoles, but not in nuclei. In cell sections the particle diameter is c. 26 nm. Infected cells (Fig. 7) contain numerous large multivesicular bodies, which appear to be derived from mitochondria, and the chloroplasts have some marginal vesicles (T. Hatta, R. I. B. Francki & C. J. Grivell, unpublished data).
Figures

Symptoms of galinsoga mosaic in naturally infected Galinsoga parviflora (healthy plant on right).

Systemic vein clearing in recently inoculated G. parviflora.

Severe systemic mosaic in chronically infected G. parviflora.
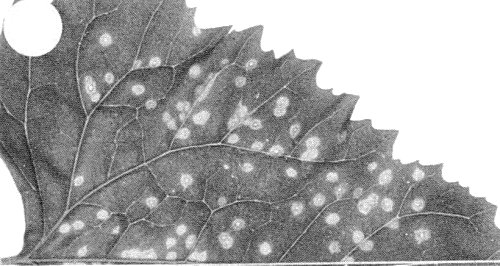
Local lesions in inoculated leaf of Chenopodium amaranticolor.
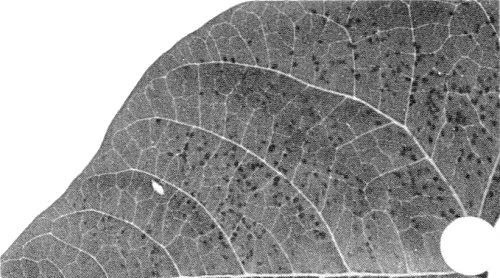
Local lesions in inoculated leaf of Phaseolus vulgaris.

Electron micrograph of particles negatively stained in uranyl acetate. Bar represents 50 nm.

Electron micrograph of a thin section of an infected leaf cell of P. vulgaris showing the characteristic multivesicular bodies and vesicles associated with the chloroplast membrane (arrows). Insert shows vesicles at the outer membrane of a multivesicular body. Sections were prepared from tissues fixed with aldehyde and treated with ribonuclease to remove ribosomal RNA, a process which leaves the virus particles unaffected (Hatta & Francki, 1981). Bars represent 200 nm.
References list for DPV: Galinsoga mosaic virus (252)
- Appiano, Pennazio & Redolfi, J. gen. Virol. 40: 277, 1978.
- Behncken, Aust. J. biol. Sci. 23: 497, 1970.
- Behncken, Aust. Pl. Path. Soc. Newsletter 1: 18, 1972.
- Hatta & Francki, J. Ultrastruct. Res. 74: 116, 1981.
- Russo & Martelli, Virology 49: 122, 1972.
- Shukla, Shanks, Teakle & Behncken, Aust. J. biol. Sci. 32: 267, 1979.
- Skotnicki &Gibbs, Australas. Pl. Path. 10: 27, 1981.