Details of DPV and References
DPV NO: 261 July 1983
Family: Betaflexiviridae
Genus: Carlavirus
Species: Hop latent virus | Acronym: HpLV
Hop latent virus
D. J. Barbara East Malling Research Station, Maidstone, Kent, England
A. N. Adams East Malling Research Station, Maidstone, Kent, England
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
Virus first clearly distinguished from hop mosaic virus by Thresh & Ormerod (1969). Virus described by Schmidt, Schmidt & Eisbein (1966), Probasco & Skotland (1978) and Adams & Barbara (1982a). These reports concern virus isolates from different countries but probably refer to the same virus.
A virus with filamentous RNA-containing particles c. 14 x 675 nm frequently occurring in hop (Humulus lupulus) in Europe, USA and Australia. Transmitted in the non-persistent manner by aphids and by mechanical inoculation. The host range is narrow.
Main Diseases
Most, if not all, hop (Humulus lupulus) cultivars show no apparent symptoms; the effects of infection on yield have not been determined. No other naturally infected host has been reported.
Geographical Distribution
Common in Europe and USA, and also occurs in Australia (Schmidt et al., 1966; Probasco & Skotland, 1978; Adams & Barbara, 1982a).
Host Range and Symptomatology
Experimentally the virus infected 11 of 60 species in 4 of 18 families: 7 species of Chenopodiaceae, 2 of Leguminosae and 1 each of Caryophyllaceae and Cannabinaceae (Probasco & Skotland, 1978; Adams & Barbara, 1982a).
- Diagnostic species
- No reliable diagnostic species is known. Seedlings derived from
open-pollinated Cluster varieties of hop developed systemic chlorotic flecking
and some systemic chlorosis and distortion following mechanical inoculation but
were variable in their response (Probasco & Skotland, 1978). No symptoms have
been reported in hops infected by grafting or by aphid inoculation.
- Propagation species
- Humulus lupulus
is suitable for maintaining cultures and as a source of virus for purification.- Assay species
- Phaseolus vulgaris
cv. Kinghorn has been used as a local lesion host. Although lesions may be induced in other cultivars their occurrence may be erratic. - Chenopodium murale. Faint chlorotic local lesions (Fig. 1) may be
produced under some conditions
but not reliably.
- Humulus lupulus has been used in aphid transmission tests.
Strains
Isolates showing minor host range differences have been reported and may represent strains but they have not been compared critically under the same conditions.
Transmission by Vectors
Transmitted in the non-persistent manner by the aphid Phorodon humuli (Adams & Barbara, 1982a). Experimental transmission from hop to hop is inefficient although spread in the field may be rapid under some circumstances (A. N. Adams, unpublished information).
Transmission through Seed
None found in hop (Adams & Barbara, 1982a).
Serology
Antisera with titres of up to 1/800 (ring interface test) were readily obtained by intramuscular and/or subcutaneous injection of rabbits with whole virus (Adams & Barbara, 1982a). The tendency of virus particles in purified preparations to aggregate makes conventional serology unreliable but the virus is readily detected in hops by ISEM or by ELISA, which has been used extensively for diagnosis.
Relationships
The properties of the virus suggest that it is a member of the carlavirus group. Serologically it is related to carnation latent, helenium S, hop mosaic, lily symptomless, nerine latent and potato M viruses; it is probably distantly related to American hop latent virus but no relationship was detected to chrysanthemum B, narcissus latent, poplar mosaic, potato S, shallot latent or red clover vein mosaic viruses (Adams & Barbara, 1982a, 1982b).
Stability in Sap
At room temperature all infectivity in sap extracts was lost in less than 30 min. The following properties were determined for leaf extracts made in the buffer described in purification method 2, and ‘stabilised’ by freezing, thawing and low speed centrifugation (Probasco & Skotland, 1978): dilution end-point, 1/100-1/1000; thermal inactivation point (10 min), 70-75°C; the virus remained infective for at least 25 days at -19°C. The thermal inactivation point of purified virus was 71°C (Probasco & Skotland, 1978).
Purification
The virus particles aggregate readily and purification may be difficult. Adams & Barbara (1982a) used two methods; the one described below (method 1) appeared to yield a purer product but was not always successful. This inconsistency did not seem to be associated with isolate or time of year. Probasco & Skotland (1978) used a different method (method 2 below); however, some English isolates did not survive freezing during this procedure.
Method 1. Extract leaves (1 g/5 ml) in phosphate-buffered saline (PBS; 0.2 g/l
potassium dihydrogen phosphate,
2.9 g/l disodium hydrogen phosphate, 8 g/l sodium chloride, 0.2 g/l potassium
chloride, pH 7.2), and centrifuge
(18,000 g for 20 min). Precipitate the virus by addition of
polyethylene glycol (PEG: M. Wt 6000)
to 50 g/l and sodium chloride to 6 g/l and sediment the precipitate by
centrifugation at 10,000 g
for 15 min. Resuspend the pellet in PBS with 0.05% Tween-20 and after one cycle
of differential centrifugation
purify the virus further by sucrose density gradient centrifugation.
Method 2. Extract leaves (1 g/6 ml) in 0.05 M sodium phosphate buffer, pH 8.0,
containing 0.2% nicotine, 0.2%
ascorbic acid and 0.5 g polyvinylpyrrolidone per gram of tissue. Filter through
cheesecloth and hold at -19°C
for 1-4 days. Thaw, centrifuge at 2000 g for 10 min and adjust
supernatant fluid to pH 5.0 with
acetic acid. Allow to stand for 30 min at 4°C and centrifuge at 2340
g for 10 min. Resuspend
the precipitate in 1/15th of the original volume of 0.01 M sodium phosphate
buffer, pH 8.0, and adjust pH to 6.8.
Centrifuge (2340 g, 30 min). Layer each 15 ml of supernatant fluid
over a 10 ml cushion composed
of 40 g/l PEG, 500 g/l sucrose, 7 g/1 sodium chloride in 0.01 M sodium phosphate,
pH 8.0, and centrifuge for 2 h
at 24,000 rev/min in a Beckman SW25.1 rotor. Resuspend pellets in 0.01 M phosphate,
pH 8.0, and further purify
by density gradient centrifugation.
Properties of Particles
Purified preparations show one component in sucrose density gradient centrifugation.
Sedimentation coefficient, s20,w: 176 S (Probasco & Skotland, 1978).
A260/A280: 1.22 (corrected for light-scattering; Adams & Barbara, 1982a; Probasco & Skotland, 1978).
Particle Structure
Particles are straight or slightly flexuous filaments c. 14 x 675 nm (Fig. 2 and Fig. 3).
Particle Composition
Nucleic acid: RNA, single-stranded, c. 5-7% of particle weight (estimated from the A260/A280 ratio). M. Wt c. 2.9 x 106 (estimated by polyacrylamide gel electrophoresis under non-denaturing conditions) (Adams & Barbara, 1982a).
Protein: c. 93-95% of particle weight (estimated from the A260/A280 ratio). M. Wt of major protein species c. 33,000, estimated by SDS/polyacrylamide gel electrophoresis. All preparations contained a variable proportion of a second species of apparent M. Wt c. 28,000, assumed to be a breakdown product or different conformational form of the major protein (Adams & Barbara, 1982a).
Relations with Cells and Tissues
No information.
Notes
At least two other viruses with similar filamentous particles occur in hop. Hop mosaic virus infects Nicotiana clevelandii systemically without inducing symptoms and causes a serious disease in some (sensitive) hop cultivars. American hop latent virus has a wider host range than either hop latent or hop mosaic viruses, infects Chenopodium quinoa systemically and is common in the USA, to which country it appears to be largely confined (Adams & Barbara, 1982a). The three viruses can occur together in hop plants that are apparently symptomless but they are distinct in host range and readily distinguishable serologically. Hop mosaic virus particles are slightly shorter than those of the other two viruses (when measured on the same specimen). Specific diagnosis in symptomless hop plants is best achieved serologically. All three viruses can be readily detected and distinguished (with no cross-reactions) by the direct form of ELISA, although significant cross-reactions were detected with indirect F(ab')2-based ELISA using globulins more concentrated than the optima for detecting homologous antigens (Adams & Barbara, 1982b).
Figures

Chenopodium murale leaves inoculated with buffer (left) or hop latent virus (right) showing diffuse chlorotic lesions in the infected leaf.

Hop latent virus particles negatively stained with 2% uranyl acetate. Bar represents 200 nm.

Mixture of particles of hop latent and hop mosaic viruses treated with antiserum to hop mosaic virus. Only the particles of the homologous virus are coated with antibodies. Negatively stained in 2% sodium phosphotungstate, pH 6.5. Bar represents 200 nm.
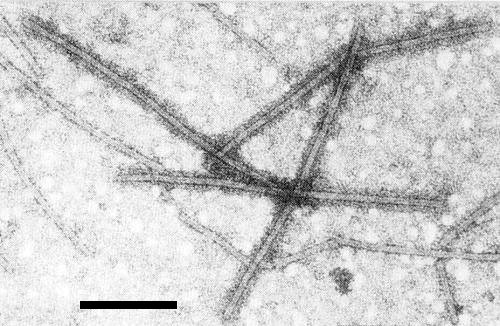
Mixture of particles of hop latent and hop mosaic viruses treated with antiserum to hop latent virus. Only the particles of the homologous virus are coated with antibodies. Negatively stained in 2% sodium phosphotungstate, pH 6.5. Bar represents 200 nm.
References list for DPV: Hop latent virus (261)
- Adams & Barbara, Ann. appl. Biol. 101: 483, 1982a.
- Adams & Barbara, Ann. appl. Biol. 101: 495, 1982b.
- Probasco & Skotland, Phytopathology 68: 277, 1978.
- Schmidt, Schmidt & Eisbein, Zentbl. Bakt. ParasitKde Abt. II 120: 461, 1966.
- Thresh & Ormerod, Rep. E. Malling Res. Stn for 1968: 41, 1969.