Details of DPV and References
DPV NO: 264 July 1983
Family: Alphaflexiviridae
Genus: Potexvirus
Species: Foxtail mosaic virus | Acronym: FoMV
Foxtail mosaic virus
Margaret N. Short John Innes Institute, Colney Lane, Norwich NR4 7UH, England
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
- Described by
Paulsen & Sill (1969) and
Paulsen & Niblett (1977).
A virus with slightly flexuous filamentous particles with a modal length of c. 500 nm, containing one coat protein species and one molecule of single-stranded RNA. Experimental host range fairly broad, many species being infected symptomlessly. No insect vector known. Reported only from the USA where the diseases caused are of no economic importance.
Main Diseases
Systemic mosaic in Setaria viridis and S. italica (Paulsen & Niblett, 1977).
Geographical Distribution
Reported only from USA.
Host Range and Symptomatology
Transmitted by manual inoculation to 56 out of 68 species of Gramineae (Paulsen & Niblett, 1977) and to 41 species in 11 dicotyledonous families (Paulsen & Niblett, 1977; Short, 1982). In many species infection is symptomless and/or confined to inoculated leaves, but the virus induces systemic mosaic in wheat, oats and barley (Fig. 1).
Diagnostic species
- Chenopodium amaranticolor.
Pin-point necrotic local lesions (Fig. 2) followed by systemic necrotic etch and then mosaic.- Gomphrena globosa. Necrotic local lesions; no systemic infection.
- Nicotiana clevelandii. Necrotic and chlorotic local lesions followed by systemic mosaic.
- Pisum sativum cv. Meteor. Necrotic local spots and rings (Fig. 3); no systemic infection.
- Tetragonia expansa. Necrotic local lesions followed by systemic mosaic.
- Nicotiana clevelandii. Necrotic and chlorotic local lesions followed by systemic mosaic.
Propagation species
- Barley is a long-lived propagation host and a good source of virus for purification.
Assay species
- Chenopodium amaranticolor
is a suitable host for local lesion assays.- Bancroft, Abou Haidar & Erickson, Virology 98: 121, 1979.
- Paulsen & Niblett, Phytopathology 67: 1346, 1977.
- Paulsen & Sill, Phytopathology 59: 1942, 1969.
- Richardson, Tollin & Bancroft, Virology 112: 34, 1981.
- Short, Rep. John Innes Inst. for 1980: 126, 1981.
- Short, Ph.D. Thesis, University of East Anglia, 1982.
Transmission by Vectors
No reports.
Transmission through Seed
Paulsen & Niblett (1977) reported 2% seed transmission in Briza maxima and 1% in Avena sativa.
Transmission by Dodder
The virus was not transmitted by Cuscuta campestris parasitic on Chenopodium quinoa, Hordeum vulgare or Nicotiana clevelandii; no virus was detected by back-inoculation from the dodder vines (Short, 1982).
Serology
The virus is strongly immunogenic: positive reactions were observed in ring interface precipitin tests with antiserum diluted to 1/1024 (Short, 1981).
Relationships
In ring interface precipitin tests, antiserum to foxtail mosaic virus reacted with purified preparations of two potexviruses, narcissus mosaic and viola mottle viruses, but not with preparations of five others: cactus X (barrel cactus strain), clover yellow mosaic, papaya mosaic, potato X or white clover mosaic viruses (Short, 1981).
Stability in Sap
Relatively stable in extracted sap, with a dilution end-point of 10-6 to 10-7 and a thermal inactivation point of 68°-70°C when heated for 10 min. Virus remained infective for 46 days in undiluted sap at room temperature, and for 105 days in buffered extract (Paulsen & Niblett, 1977).
Purification
Purified virus is readily prepared by the method devised by Bancroft, Abou Haidar & Erickson (1979) for clover yellow mosaic virus. Grind fresh or frozen leaves of Hordeum vulgare in 0.02 M sodium borate buffer, 0.01 M fresh ascorbic acid, pH 8.2, and clarify the extract with 0.5% Triton X-100. Ultracentrifuge the virus through 5 ml ‘cushions’ of 30% sucrose in 0.01 M Tris-HCl, 0.001 M EDTA, pH 8.0. Resuspend the virus in the same buffer and purify further by differential centrifugation.
Yields of 0.5 mg virus per gram leaf tissue were obtained from selected fresh leaves of Hordeum vulgare.
Properties of Particles
In linear-log sucrose gradients, purified virus particles sediment as two
u.v.-absorbing, infective components
(Paulsen & Niblett, 1977)
with sedimentation
coefficients of 122 S and 144 S and
A260/A280 = 1.2.
Both components contain a single RNA species with a sedimentation coefficient of 32 S which suggests that the 144 S component is a dimer of the 122 S component.
Particle Structure
Slightly flexuous filamentous particles with helical symmetry and a modal length of c. 500 nm when negatively stained with potassium phosphotungstate, pH 6.4 (Fig. 4). From optical diffraction studies, Richardson, Tollin & Bancroft (1981) calculated a helix pitch of 3.5 nm, with 8.8 subunits per turn; the structural parameters for foxtail mosaic virus were typical of members of the potexvirus group.
Particle Composition
Nucleic acid: Single-stranded RNA with a M. Wt of about 2.03-2.24 x 106 (Paulsen & Niblett, 1977), estimated by sedimentation with RNA standards in linear-log sucrose density gradients in 0.02 M phosphate buffer, pH 6.25. The RNA comprises about 7% of the particle weight, calculated from the data on particle structure and the RNA and coat protein subunit M. Wts.
Protein: Coat protein can be prepared by disrupting the virus in 1 M HCl. A subunit of M. Wt 21,171 was calculated from the amino acid composition and two-dimensional peptide map. The subunit polypeptide chain has 192 amino acid residues, comprising 13 lys, 1 his, 9 arg, 2 trp, 25 asp, 16 thr, 9 ser, 20 glu, 14 pro, 7 gly, 27 ala, 2 cys, 11 val, 3 met, 7 ile, 11 leu, 7 tyr, 8 phe (Short, 1981).
Relations with Cells and Tissues
Large clusters of parallel particles occur in the cytoplasm of infected cells of Nicotiana clevelandii (Fig. 5). No inclusion bodies or other virus-induced structures have been observed (K. A. Plaskitt, unpublished data).
Notes
At least 10 other viruses are readily mechanically transmissible to barley. However, these have either isometric particles (e.g. brome mosaic, cocksfoot mild mosaic, cocksfoot mottle and panicum mosaic viruses), flexuous filamentous particles 700 nm or longer (e.g. Guinea grass mosaic, sugarcane mosaic and wheat streak mosaic viruses), or rigid rod-shaped particles (e.g. barley stripe mosaic, poa semi-latent and soil-borne wheat mosaic viruses). Furthermore none of these viruses has been reported to occur naturally in Setaria spp., nor to infect Pisum sativum or Nicotiana clevelandii, and they can therefore easily be distinguished from foxtail mosaic virus by electron microscopy and host range studies. Foxtail mosaic virus has not been reported to occur naturally in barley.
Figures

Systemically infected leaves of Hordeum vulgare cv. Maris Otter.

Necrotic local lesions in Chenopodium amaranticolor.

Necrotic local spots and rings in Pisum sativum cv. Meteor.

Electron micrograph of virus particles in a leaf-dip of Nicotiana clevelandii, negatively stained in potassium phosphotungstate (courtesy G. J. Hills). Bar represents 100 nm.
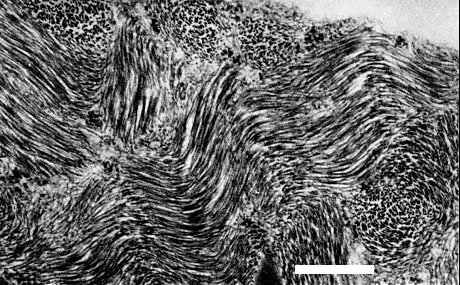
Electron micrograph of a leaf-section of Nicotiana clevelandii containing clusters of virus particles, stained in uranyl acetate (courtesy K. A. Plaskitt). Bar represents 500 nm.