Details of DPV and References
DPV NO: 273 July 1983
Family: Virgaviridae
Genus: Tobamovirus
Species: Hypochoeris mosaic virus | Acronym: HyMV
Hypochoeris mosaic virus
A. A. Brunt Glasshouse Crops Research Institute, Littlehampton, Sussex, England
R. Stace-Smith Agriculture Canada Research Station, 6660 N. W. Marine Drive, Vancouver, B.C., Canada V6T 1X2
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
Described by
Brunt & Stace-Smith (1978a,
1978b).
Possible synonym
- Cat’s ear yellow spot virus (Schmidt, 1977)
-
A virus with fragile rod-shaped particles c. 22 nm in diameter and predominant lengths of 120-140 and 220-240 nm. It is transmissible by mechanical inoculation of sap to species in eight families, but its vector is unknown. The virus occurs in eastern and western Canada, Australia and possibly also the Federal Republic of Germany.
Main Diseases
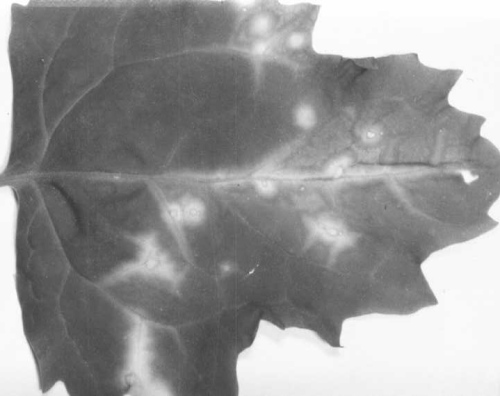
Hypochoeris radicata (cat’s ear) and Leontodon autumnalis (autumnal or fall hawkbit) are the only known natural hosts of the virus. Some leaves of infected H. radicata plants are extensively chlorotic (Fig. 1), but others are either only slightly chlorotic or symptomless (Brunt & Stace-Smith, 1978a). Young leaves of L. autumnalis develop chlorotic spots below 15°C (minimum daily temperature), but symptoms are less conspicuous at higher temperatures and are usually absent at 25°C and above.
Geographical Distribution
Prevalent in Hypochoeris radicata in scattered locations in western Canada (Brunt & Stace-Smith, 1978a), Australia (Greber & Finlay, 1981) and possibly also in the German Democratic Republic (Zschau, 1964). Locally common in Leontodon autumnalis in eastern Canada (Singh & McDonald, 1980).
Host Range and Symptomatology
In addition to Hypochoeris radicata and Leontodon autumnalis, the virus infected 16 of 56 species from 7 of 12 families (Brunt & Stace-Smith, 1978a; Singh & McDonald, 1980; Greber & Finlay, 1981).
-
Diagnostic species
- Chenopodium amaranticolor
and C. quinoa. Inoculated leaves develop chlorotic lesions (Fig. 2, Fig. 3) which sometimes extend along the veins and later become necrotic. Systemically infected leaves develop a faint chlorosis. -
Chenopodium murale. Necrotic lesions in inoculated leaves; no systemic infection.
- Cucumis sativus (cucumber). The Australian isolate induces a few chlorotic lesions in inoculated leaves, and chlorotic mottling and distortion of systemically infected leaves. Not infected by the type isolate. Susceptibility to the isolate from eastern Canada not tested.
- Gomphrena globosa. The isolate from eastern Canada induces chlorotic lesions and veinal necrosis in inoculated leaves, and the development of abnormally small systemically infected leaves with necrotic lesions. Not infected by the type isolate. Susceptibility to the Australian isolate not tested.
- Montia perfoliata. The isolate from western Canada induces severe chlorosis of systemically infected leaves, but that from Australia causes symptomless systemic infection only. Susceptibility to the isolate from eastern Canada not tested.
- Nicotiana clevelandii. Inoculated leaves develop a few grey necrotic lesions or oakleaf patterns (Fig. 4), followed by veinal necrosis and death. Systemically infected leaves are much reduced in size but are only faintly chlorotic. Plants are severely stunted.
- Tetragonia expansa. A few chlorotic lesions in inoculated leaves; no systemic infection.
- Cucumis sativus (cucumber). The Australian isolate induces a few chlorotic lesions in inoculated leaves, and chlorotic mottling and distortion of systemically infected leaves. Not infected by the type isolate. Susceptibility to the isolate from eastern Canada not tested.
-
Propagation species
- Nicotiana clevelandii
is useful for maintaining cultures, and is the best source of virus for purification.Assay species
- Chenopodium amaranticolor
and C. quinoa can be used as local lesion assay hosts.
Strains
None reported; isolates from different geographical areas have slightly different host ranges and induce different symptoms in a few common hosts (Brunt & Stace-Smith, 1978a; Singh & McDonald, 1980; Greber & Finlay, 1981).
Transmission by Vectors
Vector unknown. Not transmitted in the non-persistent manner by any of four aphid species (Aphis gossypii, Aulacorthum solani, Macrosiphum euphorbiae and Myzus persicae), or by the stubby-root nematode (Nanidorus minor) (Brunt & Stace-Smith, 1978a; Greber & Finlay, 1981).
Transmission through Seed
Not seed-borne in Hypochoeris radicata, Nicotiana clevelandii or Chenopodium quinoa (Brunt & Stace-Smith, 1978a; Greber & Finlay, 1981).
Serology
The virus is a moderately good immunogen. Antiserum obtained from a rabbit 30 days after the second of two injections, each of c. 100 µg purified virus emulsified with Freund’s complete adjuvant, had a titre of 1/1280 in tube precipitin and microprecipitin tests and 1/2560 in ring interface tests (Brunt & Stace-Smith, 1978a).
Relationships
The properties of the virus suggest that it is a possible member of the tobamovirus group, but it is serologically unrelated to tobacco mosaic virus and to the following six labile tobamoviruses: broad bean necrosis, beet necrotic yellow vein, peanut clump, potato mop-top, nicotiana velutina mosaic and wheat soil-borne mosaic. It is also unrelated to barley stripe mosaic and tobacco rattle viruses (Brunt & Stace-Smith, 1978a).
Stability in Sap
In Nicotiana clevelandii sap the virus loses infectivity after 10 min at 45-50°C, 1-2 days at 2 or 20°C and after dilution to 10-2-10-3 (Brunt & Stace-Smith, 1978a).
Purification
Yields of 4.8 mg virus/kg Nicotiana clevelandii leaf tissue are obtainable by the following procedure (Brunt & Stace-Smith, 1978a). Extract symptom-bearing leaves (1 g/3 ml) in 0.25 M borate buffer at pH 7.9 containing 0.01 M 2-mercaptoethanol, partially clarify the extract by adding chloroform (250 ml/litre), remove the aqueous phase and subject it to one cycle of differential centrifugation (15 min at 12,000 g; 75 min at 65,000 g). Resuspend the high-speed pellets in 0.05 M borate buffer at pH 7.6 (1 ml/15-30 g leaf tissue). Purify the virus further by chromatography on columns of controlled-pore glass beads (Barton, 1977), followed by chromatography on DEAE cellulose, eluting with a linear gradient of 0.1-0.5 M sodium chloride in 0.02 M neutral phosphate buffer; recover virus from the eluates by centrifugation (90 min at 65,000 g) and resuspend in 0.05 M borate buffer at pH 7.5.
Properties of Particles
Virus preparations sediment as six components which have sedimentation coefficients (s20,w) of 173, 143, 114, 71, 45 and 15 S (Brunt & Stace-Smith, 1978a).
Particle Structure
The virus has rod-shaped particles up to 420 nm long but with predominant lengths in leaf sap of 220-240 nm and 120-140 nm, and a diameter of 21-22.5 nm. The particles are fragile with fissures at irregular intervals along their lengths; many also have portions of the primary helix partially disrupted or missing (Fig. 7, Fig. 8). Fragmentation often occurs by discs separating from the particles; these discs can often be seen in end-on view or still loosely attached to particle fragments (Brunt & Stace-Smith, 1978a, 1978b).
Particle Composition
Nucleic acid: The infectivity of crude nucleic acid extracts is inactivated by RNase, but not DNase, suggesting that the nucleic acid is RNA (Brunt & Stace-Smith, 1978a).
Protein: The coat protein subunits have a M. Wt of 24.5 x 103 (estimated by electrophoresis in polyacrylamide gels containing SDS) (Brunt & Stace-Smith, 1978a).
Relations with Cells and Tissues
In infected Nicotiana clevelandii mesophyll cells, the virus particles are found scattered throughout the cytoplasm, or loosely aggregated in spherical cytoplasmic masses, or in regular arrays (Fig. 6), or associated with crystalline inclusions (Fig. 5). The crystalline inclusions (lattice spacing 9.8 nm) are enclosed in a membrane, are often rectangular in section and measure up to 2.3 x 1.5 µm; the associated particles are found occasionally within the crystals, but more frequently are adjacent to the enclosing membrane (Brunt & Stace-Smith, 1978b).
Notes
Hypochoeris mosaic virus has particles of similar morphology, size and properties to those of broad bean necrosis (Inouye & Asatani, 1968), beet necrotic yellow vein (Tamada & Baba, 1973), nicotiana velutina mosaic (Randles, Harrison & Roberts, 1976), peanut clump (Thouvenel, Dollet & Fauquet, 1976), potato mop-top (Harrison & Jones, 1970) and wheat soil-borne mosaic viruses (Brakke, Estes & Schuster, 1965), and to those of defective strains of tobacco mosaic virus (Kassanis & Woods, 1969). It also resembles some of these viruses in its intracellular location and appearance (Brunt & Stace-Smith, 1978b). Although hypochoeris mosaic virus is apparently serologically distinct, its similarities to these labile tobamoviruses suggests that it too is possibly a member of the tobamovirus group. It may also, like these labile tobamoviruses, be transmitted by a root-infecting fungus, but such transmission tests have not yet been attempted.
Figures

Naturally infected Hypochoeris radicata leaf.
Lesions in inoculated Chenopodium quinoa leaf.

Lesions in inoculated Chenopodium amaranticolor leaf.
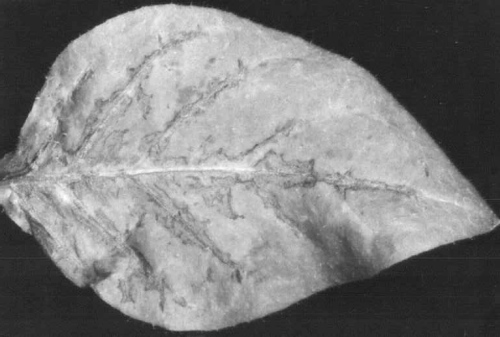
Veinal necrosis in inoculated Nicotiana clevelandii leaf.

Membrane-enclosed crystalline inclusions with embedded virus particles. Bar represents 1 µm.

Aggregated virus particles. Bar represents 100 nm.

Virus particles in sap from Nicotiana clevelandii mounted in neutral potassium phosphotungstate. Bar represents 100 nm.
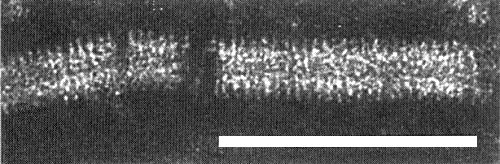
Particle at high magnification. Bar represents 100 nm.
References list for DPV: Hypochoeris mosaic virus (273)
- Barton, J. gen. Virol. 35: 77, 1977.
- Brakke, Estes & Schuster, Phytopathology 45: 70, 1965.
- Brunt & Stace-Smith, Ann. appl. Biol. 90: 205, 1978a.
- Brunt & Stace-Smith, J. gen. Virol. 39: 63, 1978b.
- Greber & Finlay, Australas. Pl. Path. 10: 30, 1981.
- Harrison & Jones, Ann. appl. Biol. 65: 393, 1970.
- Inouye & Asatani, Ann. phytopath. Soc. Japan 34: 317, 1968.
- Kassanis & Woods, Ann. appl. Biol. 64: 213, 1969.
- Randles, Harrison & Roberts, Ann. appl. Biol. 84: 193, 1976.
- Schmidt, in Pflanzliche Virologie Vol. 4: p. 517, ed. K. Schmelzer & D. Spaar, 528 pp., Berlin: Akademie-Verlag, 1977.
- Singh & McDonald, Canad. Pl. Dis. Surv. 60: 4, 1980.
- Tamada & Baba, Ann. phytopath. Soc. Japan 39: 325, 1973.
- Thouvenel, Dollet & Fauquet, Ann. appl. Biol. 84: 311, 1976.
- Zschau, NachrBl. dt. Pflschutzdienst, Berl. 18: 15, 1964.