Details of DPV and References
DPV NO: 274 July 1983
Family: Unallocated ssRNA+ viruses
Genus: Sobemovirus
Species: Southern bean mosaic virus | Acronym: SBMV
This is a revised version of DPV 57
Southern bean mosaic virus
J. H. Tremaine Agriculture Canada Research Station, 6660 N.W. Marine Drive, Vancouver, B.C., Canada V6T 1X2
R. I. Hamilton Agriculture Canada Research Station, 6660 N.W. Marine Drive, Vancouver, B.C., Canada V6T 1X2
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
Described by
Zaumeyer & Harter, 1942;
1943.
Selected synonyms
- Bean mosaic virus 4 (Rev. appl. Mycol. 21: 478)
- Southern bean mosaic virus 1 (Rev. appl. Mycol. 23: 203)
- Marmor laesiofaciens (Rev. appl. Mycol. 23: 467)
- Bean virus 4 (Rev. appl. Mycol. 31: 163)
- Bean southern mosaic virus (Rev. appl. Mycol. 37: 433)
- Phaseolusvirus laedens (Rev. appl. Mycol. 38: 677)
- Southern bean mosaic virus 1 (Rev. appl. Mycol. 23: 203)
-
A virus with isometric particles about 30 nm in diameter sedimenting as a single component and containing single-stranded RNA. The genome consists of one RNA molecule. The virus has a very restricted host range, is transmitted by beetles (Chrysomelidae), by seed and by inoculation of sap. It occurs predominantly in warm temperate and tropical regions.
Main Diseases
Causes mosaic and mottle diseases of economic importance in common bean (Phaseolus vulgaris), cowpea (Vigna unguiculata), urd bean (V. mungo) and to a lesser extent in soybean (Glycine max). Good descriptions of disease symptoms in common bean are given by Zaumeyer & Harter (1943) and Yerkes & Patino (1960); in cowpea by Shepherd & Fulton (1962); and in urd bean by Singh & Singh (1975).
Geographical Distribution
Warm temperate and tropical regions of the Americas (USA, Mexico and Costa Rica; Gamez, 1977) and Africa (Givord, 1981; Lamptey & Hamilton, 1974; Shoyinka et al., 1979). Reported also in Phaseolus vulgaris in France (Férault et al., 1969) and Brazil (Cupertino et al., 1982) and in urd bean (Vigna mungo) in India (Singh & Singh, 1975).
Host Range and Symptomatology
Host range is very narrow; with the exception of Gomphrena globosa (Givord, 1981) only species of Leguminosae are susceptible. Isolates of the bean strain (B) rarely infect cowpea, and those of the cowpea strain (C) rarely infect bean. Readily transmissible by inoculation of sap.
-
Diagnostic species
- Phaseolus vulgaris
(common bean). Strains B and M (Mexican severe strain) induce local chlorotic spots or local lesions (Fig. 1) with or without systemic spread (Fig. 3) in Bountiful, Pinto, Blue Lake, Kentucky Wonder and other varieties (Grogan & Kimble, 1964). Systemic symptoms in common bean infected by these strains range from prominent mosaic with leaf distortion (especially in younger leaves) to mild mosaic depending on variety. Strain G (Ghana) usually causes symptomless systemic infection (Lamptey & Hamilton, 1974), and strain C usually does not infect this host. - P. lunatus (lima bean). Only small-seeded varieties are susceptible
to strain B which induces
small necrotic lesions. Not reported to be susceptible to other strains.
- Glycine max (soybean). Mild systemic mottle induced by most strains depending on variety.
- Vigna radiata (mung bean). Small necrotic local lesions induced by strain G; isolates from urd bean induce systemic mosaic.
- V. unguiculata (cowpea). Strains C and G induce local lesions (Fig. 2) in cvs Georgia 21, PI 399419 and Clay (hypersensitive hosts) although strain C may occasionally infect cv. PI 399419 systemically. Systemic symptoms (Fig. 4), characterized by vein-clearing, mosaic and leaf distortion, develop in many other varieties.
- Gomphrena globosa. Systemic symptoms are induced by the Ivory Coast isolate (Givord, 1981). Not reported to be susceptible to other strains.
- Glycine max (soybean). Mild systemic mottle induced by most strains depending on variety.
-
Propagation species
- Systemically infected varieties of bean or cowpea are commonly used for maintaining virus cultures and for propagating virus for purification.
-
Assay species
- Local lesion varieties of bean (Grogan & Kimble, 1964) or cowpea (Kuhn & Brantley, 1963) are suitable.
Strains
Bean (type) strain (Strain B) (Zaumeyer & Harter, 1943). Infects most common bean varieties systemically and induces local lesions in others; can infect a limited number of other legumes but not cowpea or other legumes susceptible to the cowpea strain.
Cowpea strain (Strain C) (Shepherd & Fulton, 1962). Infects most cowpea varieties systemically; can also infect other legumes (e.g. V. unguiculata ssp. sesquipedalis) but not common bean except cv. Pinto, in which the virus causes symptomless and apparently localized infections in inoculated leaves (R. I. Hamilton, unpublished results).
Ghana strain (Strain G) (Lamptey & Hamilton, 1974). Infects many cowpea varieties systemically and also induces necrotic local lesions or systemic symptomless infection in some cultivars of common bean. The Ivory Coast isolate (Givord, 1981) is similar.
Severe bean mosaic strain or Mexican strain (Strain M) (Yerkes & Patino, 1960; Grogan & Kimble, 1964). Causes more severe symptoms (e.g. local necrosis and systemic symptoms, often with necrosis) in common bean than the type strain; also infects cowpea.
Resistance-breaking strains have been isolated from systemically infected bean cultivars resistant to strain B (Valverde & Fulton, 1982). Variants of strain B differing in electrophoretic mobility and buoyant density have also been described (Magdoff-Fairchild, 1967).
Transmission by Vectors
Transmissible by leaf beetles (Chrysomelidae), probably in a circulative manner (Walters, 1969; Fulton, Scott & Gamez, 1975). In North America, strains B and C are transmitted by Ceratoma trifurcata and Epilachna varivestis (Fulton et al., 1975); Ootheca mutabilis transmits strain C in Nigeria (Allen et al., 1981). Strain C can be transmitted by C. trifurcata for up to 19 days following acquisition access feedings (Walters & Henry, 1970) but beetle species differ in the length of time for which they continue to transmit virus without renewed access to source plants.
Transmission through Seed
Seed-borne in bean (1-5%; Zaumeyer & Harter, 1943) and more frequently in cowpea (5-40%; Shepherd & Fulton, 1962; Lamptey & Hamilton, 1974; Givord, 1981); evidence of embryo transmission in bean is equivocal (McDonald & Hamilton, 1972; Uyemoto & Grogan, 1977). Transmitted to bean seedlings if germinating seeds are in contact with infective extracts (Hamilton, 1978) or are planted in soil near roots of infected plants (Teakle & Morris, 1981). No evidence of pollen transmission to seed or to the pollinated plant but the pollen exine is contaminated (Hamilton, Leung & Nichols, 1977).
Serology
Highly immunogenic; microprecipitin titres of 1/2048-1/4096 are commonly found. Immunodiffusion in gels, including the use of antisera to sodium dodecyl sulphate-treated virus (Purcifull, Christie & Lima, 1981), immunosorbent electron microscopy and ELISA are applicable for detecting virus in tissue extracts.
Relationships
The physical and chemical properties of southern bean mosaic virus particles are similar to those of other members of the sobemovirus group such as turnip rosette, blueberry shoestring, cocksfoot mottle, rice yellow mottle, sowbane mosaic (Matthews, 1982) and lucerne transient streak viruses (Forster & Jones, 1980) but the virus is not serologically related to any other members except for a possible distant relationship of the Ivory Coast isolate to sowbane mosaic virus (Givord, 1981). All strains of southern bean mosaic virus are serologically related to each other but strains B and M can be distinguished from strains C and G by immunodiffusion in gels (Grogan & Kimble, 1964; Lamptey & Hamilton, 1974; R. I. Hamilton, unpublished results).
Stability in Sap
In bean or cowpea sap: infectivity dilution end-point, 10-5-10-8; thermal inactivation point (10 min) 90-95°C; longevity (18-22°C) 20-165 days (Zaumeyer & Harter, 1942; Yerkes & Patino, 1960; Shepherd & Fulton, 1962; Lamptey & Hamilton, 1974; Givord, 1981).
Purification
Easily purified in high yield (1 mg/g infected fresh leaf tissue); yields obtained with tissue frozen at -10°C for longer than 1 month are much lower.
Method 1. (Ghabrial, Shepherd & Grogan, 1967, as modified by Johnson et al., 1974). Extract the infected tissue 3 weeks after inoculation in 0.5 M sodium citrate (pH 7.5) containing 0.1% 2-mercaptoethanol and clarify the extract by low speed centrifugation. Adjust the supernatant fluid to pH 4.5 with 10% acetic acid and after 1 h centrifuge the material at low speed. Adjust the supernatant fluid to pH 6 with 10% NaOH and precipitate the virus by adding polyethylene glycol, M. Wt 6000 (PEG), to 8% (w/v) and NaCl to 0.1% (w/v). Give the preparation four to five cycles of differential centrifiigation and use 0.05 M phosphate (pH 7.0) as a suspending medium. Adjust the preparation to pH 4.5 between the second and third cycles.
Method 2. (Tremaine, Ronald & Kelly, 1981). Homogenize freshly harvested infected leaves in 0.2 M sodium acetate buffer (pH 5.0) containing 0.02 M sodium diethyldithiocarbamate and 0.1% 2-mercaptoethanol. Adjust the extract to pH 4.8 with 10% acetic acid, and leave at 5°C for 4 h. Clarify the extract by low speed centrifugation and precipitate the virus with PEG at 8% (w/v). Suspend the pellet from low speed centrifugation in 0.1 M sodium acetate buffer (pH 5.0) and give the preparation two cycles of differential centrifugation. High speed pellets turn white and become insoluble when suspended at pH 5.0 but the B strain is very soluble when suspended in 0.1 M sodium phosphate buffer (pH 7.0). The C strain is less soluble in this buffer (8 mg/ml) but more soluble after addition of 1 M NaCl. Particles suspended at pH 7.0 are stable and soluble after dialysis or adjustment to pH 5.0.
The virus can also be purified with the aid of organic solvents (Hull, 1977a). Strain B crystallizes on dialysis against distilled water (Miller & Price, 1946b) or in solutions of MgSO4 or (NH4)2SO4 (Price, 1946). Crystals of strain C useful in small angle X-ray diffraction are formed when samples at 20 mg/ml are treated with 0.95 M (NH4)2SO4 (Johnson et al., 1974).
Properties of Particles
The virus particles are stable between pH 2.5 and 9.5 (Sehgal, 1980). The particles swell on treatment with EDTA under slightly alkaline conditions (Wells & Sisler, 1969) and dissociate into RNA and protein with the further addition of 1 M NaCl (Hull, 1977b). The particles are stabilized by divalent ion-dependent and pH-dependent protein-protein interactions and by RNA-protein interactions (Hull, 1977b). Stable virus particles can be reconstituted from B strain protein and either B strain RNA or sowbane mosaic virus RNA (Tremaine & Ronald, 1977).
Sedimentation coefficient: The particles sediment as a single component with s20,w (svedbergs) = 115 S at infinite dilution.
Particle weight (daltons): 6.6 x 106 (Miller & Price, 1946a); 6.1 x 106 (Lauffer, Taylor & Wunder, 1952); 6.5 x 106 (Yphantis, 1964).
Diffusion coefficient (D20,w x 10-7cm2/sec): 1.34 (Miller & Price, 1946a).
Partial specific volume: 0.696-0.700 (Miller & Price, 1946a; Lauffer et al., 1952).
Electrophoretic mobility: mobility varies with the strain (Hartmann & Lauffer, 1953; Magdoff-Fairchild, 1967; Tremaine & Wright, 1967).
Absorption coefficient A260(0.1%, 1 cm): 5.85.
A260/A280: 1.6.
Buoyant density (g/cm3): 1.360 in CsCl; 1.402 in CsBr; 1.282 in metrizamide; two banding zones in Cs2SO4 gradients, single band in heavy zone (1.32) and one to four bands in light zone (1.28-1.305) (Hull, 1977a); 1.26 in sucrose (Lauffer et al., 1952).
Particle Structure
Particles are isometric, approximately 30 nm in diameter (Fig. 5) and have a T = 3 structure with 180 protein subunits. Negatively stained particles do not show distinct capsomeric structures in the electron microscope because the high protein density at the surface of the particle masks the differentiation of morphological features. Pentamer-hexamer clustering and an additional trimer clustering has been detected in the C strain by X-ray diffraction at 22.5 Å resolution (Johnson et al., 1976). Higher resolution X-ray diffraction studies at 2.8 Å (Abad-Zapatero et al., 1980) showed the protein is an eight-stranded, antiparallel beta barrel with the N-terminus forming an additional partially ordered arm in the interior of the virus particle. There are three conformations of the protein subunit (A, B, and C) within one icosahedral asymmetric unit. The exact three-dimensional positioning of residues 42 to 260 of the C subunit including those residues in contact with the RNA, with adjacent subunits or with the exterior are known (Hermodson et al., 1982). The Ca2+ ion is located on the quasi-threefold axes between subunits A, B, and C (Abdel-Meguid et al., 1981). The structure of the protein subunit of southern bean mosaic virus is very similar to that of tomato bushy stunt virus (Harrison et al., 1978) and the satellite virus of tobacco necrosis virus (Liljas et al., 1982).
Particle Composition
Nucleic acid: Single-stranded RNA, about 21% of the particle weight (Miller & Price, 1946a). RNA is best prepared from particles dissociated in SDS, EDTA and Tris buffer (pH 8) and extracted by adding two vol of a 1:1 vol/vol mixture of chloroform and phenol (Salerno-Rife, Rutgers & Kaesberg, 1980). The genome is contained in a single RNA molecule of M. Wt 1.4 x 106 and sedimentation coefficient of 24.9 S, becoming 14.9 S after treatment with formaldehyde (Diener, 1965; Kaper & Waterworth, 1973). Molar percentage of nucleotides G27; A22-24; C22-24; U27 (Ghabrial et al., 1967). RNA preparations also contain two prominent subgenomic RNA components as well as heterogeneous RNA (Rutgers, Salerno-Rife & Kaesberg, 1980).
Protein: Coat protein can be isolated by first swelling the virus
(Hsu, Sehgal & Pickett, 1976)
at 10-20 mg/ml in 0.1 M sodium phosphate buffer, 0.01 M EDTA (pH. 8.0) for at least 1 h at
0°C and then adding an equal volume of 4 M LiCl and freezing overnight at -20°C.
After thawing,
the RNA precipitate is removed by low speed centrifugation and the protein-containing
supernatant
fluid dialysed against 0.01 M phosphate buffer (pH 7.5) containing 0.01 M EDTA to
remove LiCl.
A260/A280 = 0. 5 and
A280nm (0.1%, 1 cm) = 1.3
(Erickson & Rossmann, 1982).
The particles contain a single protein species comprising 79% of the particle weight; the amino acid composition differs for the B, C, and M strains (Tremaine, 1966; Ghabrial et al., 1967). The coat protein of strain C consists of 260 residues of M. Wt 28,218 with a known sequence (Hermodson et al., 1982). The amino acid sequences of residues 174 to 260 of strains B and C have been determined from the nucleic acid sequences (Mang, Ghosh & Kaesberg, 1982). The cyanogen bromide peptide electrophoresis patterns of strains C and G are similar, as are those of strains B and M. The composition of the amino terminal CNBr peptide of strain B is known (Tremaine et al., 1981).
Genome Properties
The single RNA species has a covalently linked 5' terminal protein; no polyadenylated terminal region or tRNA like structure has been detected (Ghosh et al., 1979). The proteins at the 5' termini of the RNA molecules of strains B and C have M. Wt of 12,300 and 10,000 respectively and are required for infectivity (Veerisetty & Sehgal, 1980; Mang et al., 1982). The sequences of 400 bases of the 3' termini of the RNA molecules of strains B and C have been determined; there is no obvious sequence homology in the non-coding regions of the 3' termini of the RNA from strains B (129 nucleotides) and C (135 nucleotides). The remainder of the sequenced portion is the coat protein cistron, where there is extensive base and amino acid sequence homology (Mang et al., 1982).
Translation of unfractioned RNA of strain B in the wheat embryo or rabbit reticulocyte systems results in two related products, P1 and P2, and two distinctive products, P3 (the coat protein) and P4 (Salerno-Rife et al., 1980). The proteins have M. Wt of 105,000, 75,000, 29,000 and 14,000 respectively. P4 is not related to the genome-linked protein. Fractionation of the RNA by density gradient centrifugation yields the full length RNA, a 0.7 to 0.9 x 106 component and a 0.3 to 0.4 x 106 component as well as heterogeneous populations of intermediate and smaller size (Rutgers et al., 1980; Mang et al., 1982). Translation of the full-sized RNA yields P1, P2 and P4; the 0.7 to 0.9 x 106 RNA yields P2 and P4; the 0.3 to 0.4 x 106 RNA yields P3; and most of the intermediate fractions yield P4. Translation of the RNA from strain C is essentially identical but the proteins differ somewhat in size (100,000, 70,000, 30,000 and 20,000).
Relations with Cells and Tissues
Virus particles of both strains B and C are found in cytoplasm and nuclei of infected mesophyll cells (De Zoeten & Gaard, 1969); crystallization of virus particles in mesophyll cells is frequent in cowpea 5-19 days after inoculation (Fig. 6), rare in bean (Weintraub & Ragetli, 1970). Mitochondria may contain dense bodies and extrusions of chloroplasts may also occur (Weintraub & Ragetli, 1970). Removal of ribosomes by in situ RNAse treatment (Hatta & Francki, 1981) clearly revealed randomly distributed virus particles in the cytoplasm.
Notes
The virus can be distinguished from other legume-infecting viruses by its morphology, sedimentation rate, serology and narrow host range.
Figures

Local lesions induced in primary leaf of Phaseolus vulgaris (cv. Pinto) inoculated with strain B.

Local lesions induced in primary leaf of Vigna unguiculata (cv. Georgia 21) inoculated with strain C.

Systemic mosaic induced in leaf of P. vulgaris (cv. Black Turtle Soup) infected with strain M.

Vein clearing and distortion in systemically infected leaf of V. unguiculata (cv. California Blackeye) infected with strain C.
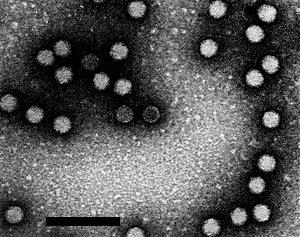
Virus particles from a purified preparation mounted in 2% uranyl acetate. Bar represents 100 nm.

Section of leaf mesophyll cell of V. unguiculata infected with strain C showing groups of virus particles in crystalline array. (Courtesy, M. Weintraub.) Bar represents 1 µm.
References list for DPV: Southern bean mosaic virus (274)
- Abad-Zapatero, Abdel-Meguid, Johnson, Leslie, Rayment, Rossmann, Suck & Tsukihara, Nature, Lond. 286: 33, 1980.
- Abdel-Meguid, Yamane, Fukuyama & Rossmann, Virology 114: 81, 1981.
- Allen, Anna-Nyako, Ochieng & Ratinam, Trop. Agric. Trin. 58: 171, 1981.
- Cupertino, Lin, Kitajima & Costa, Pl. Dis. 66: 742, 1982.
- De Zoeten & Gaard, J. Cell Biol. 40: 814, 1969.
- Diener, Virology 27: 425, 1965.
- Erickson & Rossmann, Virology 116: 128, 1982.
- Férault, Spire, Bannerot, Bertrandy & Le Tan, Annls Phytopath. 1: 619, 1969.
- Forster & Jones, CMI/AAB Descriptions of Plant Viruses, 224, 4 pp., 1980.
- Fulton, Scott & Gamez, in Tropical Diseases of Legumes, eds. J. Bird & K. Maramorosch, New York: Academic Press, p. 123, 1975.
- Gamez, Fitopatologia, Bogota 12: 24, 1977.
- Ghabrial, Shepherd & Grogan, Virology 33: 17, 1967.
- Ghosh, Dasgupta, Salerno-Rife, Rutgers & Kaesberg, Nucl. Acids Res. 7: 2137, 1979.
- Givord, Pl. Dis. 65: 755, 1981.
- Grogan & Kimble, Phytopathology 54: 75, 1964.
- Hamilton, Abstr. 4th int. Congr. Virol: 647, 1978.
- Hamilton, Leung & Nichols, Phytopathology 67: 395, 1977.
- Harrison, Olsen, Schutt, Winkler & Bricogne, Nature, Lond. 276: 368, 1978.
- Hartmann & Lauffer, J. Am. chem. Soc. 75: 6205, 1953.
- Hatta & Francki, J. Ultrastruct. Res. 74: 1, 1981.
- Hermodson, Abad-Zapatero, Abdel-Meguid, Pundak, Rossmann & Tremaine, Virology 119: 133, 1982.
- Hsu, Sehgal & Pickett, Virology 69: 587, 1976.
- Hull, Virology 79: 50, 1977a.
- Hull, Virology 79: 58, 1977b.
- Johnson, Rossmann, Smiley & Wagner, J. Ultrastruct. Res. 46: 441, 1974.
- Johnson, Akimoto, Suck, Rayment & Rossmann, Virology 75: 394, 1976.
- Kaper & Waterworth, Virology 51: 183, 1973.
- Kuhn & Brantley, Pl. Dis. Reptr 47: 1094, 1963.
- Lamptey & Hamilton, Phytopathology 64: 1110, 1974.
- Lauffer, Taylor & Wunder, Archs Biochem. Biophys. 40: 453, 1952.
- Liljas, Unge, Jones, Fridborg, Lövgren, Skoglund & Strandberg, J. molec. Biol.159: 93, 1982.
- McDonald & Hamilton, Phytopathology 62: 387, 1972.
- Magdoff-Fairchild, Virology 31: 142, 1967.
- Mang, Ghosh & Kaesberg, Virology 116: 264, 1982.
- Matthews, Intervirology 17: 1, 1982.
- Miller & Price, Archs Biochem. 10: 467, 1946a.
- Miller & Price, Archs Biochem. 11: 329, 1946b.
- Price, Am. J. Bot. 33: 45, 1946.
- Purcifull, Christie & Lima, Phytopathology 71: 1221, 1981.
- Rutgers, Salerno-Rife & Kaesberg, Virology 104: 506, 1980.
- Salerno-Rife, Rutgers & Kaesberg, J. Virol. 34: 51, 1980.
- Sehgal, Phytopathology 70: 342, 1980.
- Shepherd & Fulton, Phytopathology 52: 489, 1962.
- Shoyinka, Bozarth, Reese & Okusanya, Turrialba 29: 111, 1979.
- Singh & Singh, Phytopath. Mediterranea 14: 55, 1975.
- Teakle & Morris, Pl. Dis. 65: 599, 1981.
- Tremaine, Virology 30: 348, 1966.
- Tremaine & Ronald, Can. J. Bot. 55: 2274, 1977.
- Tremaine & Wright, Virology 31: 481, 1967.
- Tremaine, Ronald & Kelly, Can. J. Microbiol. 27: 654, 1981.
- Uyemoto & Grogan, Phytopathology 67: 1190, 1977.
- Valverde & Fulton, Phytopathology 72: 1265, 1982.
- Veerisetty & Sehgal, Phytopathology 70: 282, 1980.
- Walters, Adv. Virus Res. 15: 339, 1969.
- Walters & Henry, Phytopathology 60: 177, 1970.
- Weintraub & Ragetli, J. Ultrastruct. Res. 32: 167, 1970.
- Wells & Sisler, Virology 37: 227, 1969.
- Yerkes & Patino, Phytopathology 50: 334, 1960.
- Yphantis, Biochemistry 3: 297, 1964.
- Zaumeyer & Harter, Phytopathology 32: 438, 1942.
- Zaumeyer & Harter, J. agric. Res. 67: 305, 1943.