Details of DPV and References
DPV NO: 277 July 1984
Family: Secoviridae
Genus: Comovirus
Species: Ullucus virus C | Acronym: UVC
Ullucus virus C
A. A. Brunt Glasshouse Crops Research Institute, Littlehampton, West Sussex, England
Sue Phillips Glasshouse Crops Research Institute, Littlehampton, West Sussex, England
R. J. Barton Glasshouse Crops Research Institute, Littlehampton, West Sussex, England
R. A. C. Jones Harpenden Laboratory, Hatching Green, Harpenden, Herts, England
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
Described by
Brunt, Barton, Phillips & Jones (1982b).
A virus prevalent in Ullucus tuberosus crops in the Andean highlands of South America. It is readily transmitted by mechanical inoculation of sap to a narrow range of hosts, and has isometric particles c. 28 nm in diameter with properties typical of comoviruses.
Main Diseases
In Ullucus tuberosus (Basellaceae), its only known natural host, the virus usually occurs together with two or three other viruses (Brunt, Phillips, Jones & Kenten, 1982a). Such plants are stunted and have some chlorotic leaves, but plants infected experimentally with ullucus virus C alone remain symptomless (Brunt et al., 1982b).
Geographical Distribution
Reported only from Bolivia and Peru (Brunt et al., 1982a).
Host Range and Symptomatology
Transmitted readily by inoculation of sap to Ullucus tuberosus and to 5 of 26 species from 3 of 8 families (Brunt et al., 1982b).
-
Diagnostic species
- Chenopodium amaranticolor
and C. quinoa. Faint chlorotic lesions in inoculated leaves after 7-10 days (Fig. 1), followed by chlorosis of systemically infected leaves (Fig. 4). - Chenopodium murale. A few lesions in inoculated leaves after 14-21
days
(Fig. 2).
No systemic infection.
- Tetragonia expansa. Dark green ring lesions in inoculated leaves after c. 21 days (Fig. 3). No systemic infection.
-
Propagation species
- Chenopodium quinoa
is a convenient source of virus for purification and is a good host for maintaining cultures.Assay species No reliable local lesion assay host is known; lesion formation in Chenopodium murale and Tetragonia expansa is erratic, and lesions in C. amaranticolor and C. quinoa are often inconspicuous. Better infectivity assays are made by recording the proportion of inoculated Chenopodium quinoa seedlings that are infected systemically.
Strains
None reported.
Transmission by Vectors
Vector unknown. The virus is not transmitted in the non-persistent manner by the aphids Aphis gossypii or Myzus persicae (Brunt et al., 1982b); beetle-transmission has not been attempted.
Transmission through Seed
Not seed-borne in Chenopodium quinoa (Brunt et al., 1982b).
Serology
The virus is a good immunogen. Antiserum, obtained from a rabbit injected intramuscularly (two injections, 2 weeks apart, each of c. 2 mg of virus emulsified in Freund’s complete adjuvant) reacted to a homologous titre of 1/4096 in tube precipitin and agar double-diffusion tests.
Relationships
Ullucus virus C is serologically distantly related to cowpea mosaic virus, but it showed no relationship to eleven other recognised comoviruses (Andean potato mottle, bean pod mottle, bean rugose mosaic, broad bean stain, cowpea severe mosaic, echtes Ackerbohnemosaik, glycine mosaic, quail pea mosaic, radish mosaic, red clover mottle and squash mosaic viruses) or to broad bean wilt virus (Brunt et al., 1982b).
Stability in Sap
Sap from systemically infected Chenopodium quinoa leaves is usually infective after dilution to 10-7 but not 10-8, and after heating for 10 min at 70 but not 75°C. Infectivity of sap stored at c. 20°C is lost after 8-16 weeks, but is retained in desiccated leaves for at least 18 months.
Purification
The virus is best purified from infected Chenopodium quinoa plants, the following procedure yielding up to 40 mg virus per kg leaf tissue (Brunt et al., 1982a). Homogenise infected leaves (1 g/3 ml) in 0.07 M phosphate buffer (pH 7.6) containing 0.14 M mercaptoacetate, and clarify by adding n-butanol to 8.5% (v/v). After 30 min, subject the mixture to one cycle of differential centrifugation (20 min at 12,000 g; 90 min at 65,000 g) and resuspend the final pellets in 0.03 M neutral phosphate buffer (1 ml/20 g leaf material). Purify preparations further either by permeation chromatography on columns of controlled-pore glass beads (700 Å; 200 mesh) or, especially if separation of components is required, by rate-zonal sucrose density gradient centrifugation, and reconcentrate the virus by high speed centrifugation (120 min at 65,000 g).
Properties of Particles
The particles sediment as three components: apparently empty protein shells (T), and two kinds of nucleoprotein containing different amounts of RNA (M and B).
Sedimentation coefficients (s°20,w): 51 S (T), 95 S (M) and 116 S (B).
Buoyant density in CsCl (g cm-3): 1.404 (M); 1.463 (B).
A260/A280: 1.58 (M) and 1.65 (B); A260/A240: 1.29 (M) and 1.45 (B).
Particle Structure
Particles are isometric, c. 28 nm in diameter and slightly hexagonal in outline. When unfractionated preparations are mounted in 2% neutral potassium phosphotungstate, many particles are unpenetrated, but others are penetrated either partially or completely by the electron-dense stain (Fig. 5).
Particle Composition
Nucleic acid. Single-stranded RNA; c. 30% and 38% by weight of M and B particles, respectively (estimated from sedimentation coefficients and buoyant densities); M particles contain RNA of M. Wt 1.45 x 106 and B particles contain RNA of M. Wt 2.2 x 106 (estimated by polyacrylamide gel electrophoresis under non-denaturing conditions).
Protein: All particles contain equimolar amounts of two polypeptides with M. Wts of 21.1 x 103 and 45.1 x 103 (estimated by SDS/polyacrylamide gel electrophoresis). Particles contain a total of 3.95 x 106 daltons of protein.
Relations with Cells and Tissues
No information.
Notes
Ullucus tuberosus has been grown as a food crop in the Andean highlands of Peru, Argentina and Bolivia for well over 1000 years (Hodge, 1951). It is still an important food crop in some South American countries, especially in Peru where production in 1973, the last year for which statistics are available, exceeded 66,000 tonnes (Kay, 1973). U. tuberosus crops are grown in small terraced fields at altitudes of 2500 to 4250 metres usually near, or interplanted with, other vegetatively propagated tuber crops such as potato (Solanum tuberosum), isañu or mashua (Tropaeolum tuberosum), oca (Oxalis tuberosa) and arracacha (Arracacia xanthorrhiza). Ullucus virus C is apparently distinct from other manually transmissible viruses with isometric particles infecting these crops, such as Andean potato latent, Andean potato mottle, arracacha A and arracacha B viruses, and potato black ringspot virus and its close relation, the potato calico strain of tobacco ringspot virus. However, further surveys are necessary to determine whether it occurs naturally in other crops or weed species, especially Chenopodium quinoa, which is very susceptible to experimental infection and is grown in the Andes as a grain crop.
Virus-free Ullucus tuberosus plants can be obtained from naturally-infected plants by meristem-tip culture (Stone, 1982).
Figures

Lesions in inoculated Chenopodium amaranticolor leaf.

Lesions in inoculated C. murale leaf.
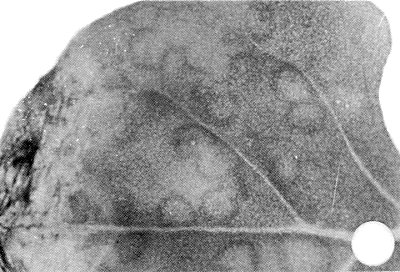
Lesions in inoculated Tetragonia expansa leaf.

Mild chlorosis in systemically infected C. amaranticolor leaf (right); healthy leaf (left).

Virus particles mounted in neutral potassium phosphotungstate. Bar represents 50 nm.
References list for DPV: Ullucus virus C (277)
- Brunt, Phillips, Jones & Kenten, Ann. appl. Biol. 101: 65, 1982a.
- Brunt, Barton, Phillips & Jones, Ann. appl. Biol. 101: 73, 1982b.
- Hodge, Econ. Bot. 5: 185, 1951.
- Kay, in Crop and Product Digest No. 2: p. 7, London: The Tropical Products Institute, 1973.
- Stone, Ann. appl. Biol. 101: 79, 1982.